DOI:
10.1590/1089-6891v18e-39566
ZOOTECNIA
MODELOS DE REGRESSÃO ALEATÓRIA PARA CARACTERÍSTICA DE
CRESCIMENTO EM BOVINOS DA RAÇA GUZERÁ
RANDOM REGRESSION MODELS FOR GROWTH TRAIT IN GUZERA CATTLE
Jorge Luis Ferreira1*
Tiago Bresolin2
Fernando Brito Lopes3
José Américo Soares Garcia4
Leandro Lopes Nepomuceno1
Aline Beatriz Schmidt1
Raysildo Barbosa Lobo5
1Universidade Federal do Tocantins, Araguaína,
TO, Brasil
2Faculdade de Ciências Agrárias e Veterinárias da UNESP,
Jaboticabal, SP, Brasil
3College of Agricultural & Life Sciences, University of
Wisconsin, Madison, Wiscosin, USA.
4Universidade de Brasília, Brasília, DF, Brasil.
5Associação Nacional de Criadores e Pesquisadores, Ribeirão
Preto, SP, Brasil.
*Autor para correspondência - jlferreira@mail.uft.edu.br
Resumo
O objetivo do presente trabalho foi estimar componentes de (co)variâncias
e parâmetros genéticos para características de pesos do nascimento ao
sobreano (550 dias de idade) para bovinos da raça Guzerá, por meio de
modelos de regressão aleatória. Os dados utilizados são de animais (machos
e fêmeas) nascidos entre 1993 e 2011 e pertencentes a dez fazendas de sete
estados brasileiros. O modelo de melhor ajuste considerou os efeitos
genéticos aditivos diretos e maternos e os de ambiente permanente do
animal ajustado usando polinômios ortogonais de Legendre de quarta,
terceira e segunda ordem, respectivamente. As maiores estimativas de
herdabilidade diretas foram observadas para os pesos ao nascimento (0,88)
e ao sobreano (0,70). As correlações genéticas foram de moderada a alta
magnitude, mantendo-se elevada mesmo com o aumento da distância entre as
idades.
Palavras-chave: bovinos de corte; correlações genéticas;
herdabilidade; polinômios ortogonais de Legendre.
Abstract
The aim of this study was to estimate (co)variance components and genetic
parameters from birth to yearling weight (550 days of age) in Guzerá
cattle, using random regression models. Data set from animals (male and
female) born between 1993 and 2011 belonging to ten farms in seven
Brazilian States were used. The best-fit model considering direct and
maternal genetic effect and permanent environmental animal effect were
adjusted by fourth, third, and second order, respectively, using
orthogonal polynomials of Legendre. The higher estimated values of direct
heritability were observed in birth (0.88) and yearling weight (0.70). The
genetic correlations were moderate to high magnitude, remaining up even
with the increase in the ages gap.
Key words: beef cattle; genetic correlations;
heritability; orthogonal polynomials of Legendre.
Recebido em: 27 janeiro de 2016
Aceito em: 14 dezembro de 2016
Introdução
Os programas de melhoramento genético de bovinos no Brasil
têm preconizado a seleção para características de crescimento, como ganho
de peso ou pesos calculados para idades-padrão. Essas características são
de fácil mensuração e apresentam herdabilidade de moderada a alta
magnitude e, portanto, são amplamente utilizadas como critérios de
seleção, resultando em maior progresso genético(1,2). No
entanto, algumas características como tamanho corporal ou de crescimento,
quando avaliadas no mesmo indivíduo em várias idades, comumente apresentam
medidas chamadas de longitudinais(3,4). Essas medidas
normalmente são utilizadas em modelos de características múltiplas, os
quais podem ser super parametrizados ao se utilizarem medidas em várias
idades. Além disso, a matriz de covariâncias não é estruturada e, para que
se possam fazer inferências sobre medidas no tempo, é importante modelar
adequadamente a estrutura de covariâncias(5,6), levando em
consideração o efeito temporal que atua e conecta cada mensuração.
Nos últimos anos tem-se proposto a utilização de modelos de regressão
aleatória para se analisarem as características observadas várias vezes ao
longo da vida do animal(4,7-8). Medidas longitudinais de peso
que se alteram com a idade podem ser representadas por uma trajetória em
função do tempo. Assim, um determinando valor em cada uma das possíveis
idades, em que registros repetidos da mesma característica e medidos em
idades diferentes, passa a representar diferentes características. Tais
trajetórias são denominadas de características de “dimensão infinita”(3,9,10),
desenvolvida por meio de um modelo matemático de regressão sobre o tempo,
para que se possa estimar parâmetros e funções da curva.
Dessa forma, o objetivo deste estudo foi estimar componentes de
(co)variâncias e parâmetros genéticos para peso do nascimento ao sobreano
(550 dias de idade), utilizando-se modelos de regressão aleatória com
diferentes ordens de ajuste para os efeitos genéticos aditivos diretos e
maternos e os de ambiente permanente de animal para bovinos da raça
Guzerá.
Materiais e Métodos
Foram utilizadas medidas de peso e idade de bovinos da raça
Guzerá, provenientes da Associação Nacional dos Criadores e Pesquisadores
– ANCP, mensuradas do nascimento aos 550 dias de idade. Os animais são
oriundos de fazendas localizadas nos estados do São Paulo (SP), Mato
Grosso do Sul (MS), Mato Grosso (MT), Goiás (GO), Tocantins (TO), Pará
(PA) e Rondônia (RO), nascidos entre 1993 e 2011. No conjunto de dados
foram mantidos apenas animais com genealogia conhecida e registro de peso
com a informação da idade. Foram consideradas três estações de nascimento
(janeiro a abril; maio a agosto; setembro a dezembro). Os grupos de
contemporâneos (GC) reuniram animais nascidos no mesmo ano e estação, do
mesmo sexo, pertencentes à mesma fazenda e criados no mesmo grupo de
manejo.
Para a consistência do arquivo de dados (Tabela 1) foram realizadas
algumas restrições por meio de rotinas desenvolvidas em linguagem do SAS(11),
considerando-se apenas animais com mais de três mensurações de peso, GC
com mais de quatro animais e touros com mais de três progênies. Além
disso, excluíram-se registros de peso fora dos intervalos de mais ou menos
três desvios-padrão, dado pela média do GC. A conexidade entre os GC foi
observada por meio do software AMC(12), com base no número
total de laços genéticos, considerando-se no mínimo dez laços genéticos.

Para a estimação dos componentes de (co)variâncias foram realizadas
análises uni-característica, pelo método da máxima verossimilhança
restrita livre de derivadas, através do modelo animal, ajustando-se
polinômios ortogonais de Legendre variando de segunda a quarta ordem para
modelar os efeitos genéticos aditivos direto e maternos e de ambiente
permanente do animal. A não inclusão do efeito de ambiente permanente
materno se deve à proporção de apenas 1,6 filhos por vaca. As análises
foram realizadas com o software WOMBAT(13) e o algoritmo AIREML
com o critério de convergência padrão menor que 10-8.
Os modelos consideraram os efeitos aleatórios genéticos aditivos diretos e
materno e de ambiente permanente do animal como fixos e o efeito de GC,
com as idades do animal à pesagem e da vaca ao parto (efeitos linear e
quadrático) como covariáveis. A curva média da população foi modelada
considerando-se o polinômio ortogonal de Legendre de quarta ordem e, para
agrupar classes de variâncias residuais semelhantes, formaram-se oito
classes, de acordo com a idade em dias (1, 2-45, 46-90, 91-140, 141-180,
181-230, 231-320, 321-550).
O modelo geral de regressão aleatória utilizado foi:
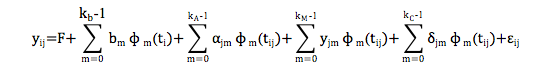
Em que: yij é a iésima medida referente ao jésimo
animal; F é o conjunto de efeitos fixos; bm é o coeficiente de
regressão fixo para modelar a curva média da população; ɸm (ti)
é a função de regressão que descreve a curva média da população de acordo
com a idade do animal; ɸm (tij) é a função de regressão que
descreve as trajetórias de cada indivíduo j, de acordo com a idade (ti),
para os efeitos aleatórios genéticos aditivos direto e materno e de
ambiente permanente do animal; αjm, yjm , δjm,
são os coeficientes de regressão genéticos aditivos direto e materno e de
ambiente permanente de animal, respectivamente, para cada animal; kb,
kA, kM e kC são as ordens dos polinômios
a serem ajustados; εij é o erro aleatório associado a cada
idade i do animal j. Na forma matricial, o modelo fica expresso como:
y = Xb + Z1α + Z2ϒ + W1δ
+ ε
Em que: y é o vetor de observações; b é o vetor de efeitos
fixos; α é o vetor aleatório dos coeficientes de regressão genéticos
aditivos diretos; ϒ é o vetor aleatório dos coeficientes de regressão
genéticos maternos; δ é o vetor aleatório dos coeficientes de regressão de
ambiente permanente do animal; X, Z1, Z2 e W1
são as matrizes de incidência correspondentes às observações, para efeitos
fixos, aleatórios genéticos aditivos direto e materno e de ambiente
permanente de animal, respectivamente; ε é o vetor de resíduos.
As pressuposições em relação aos componentes são:

Em que KA, KM e KC são as
matrizes de (co)variâncias entre os coeficientes de regressão aleatórios
para os efeitos genéticos aditivos direto e materno e de ambiente
permanente do animal, respectivamente; A é a matriz de parentesco; I é a
matriz identidade; Na é o número de animais com pesagens; ⨂ é o
produto de Kroneker entre matrizes; R é a matriz de variâncias residuais.
Para comparar os diferentes modelos de regressão aleatória testados, foram
utilizados os critérios de informação de Akaike (AIC = - 2 logL + 2p) e
Bayesiano de Schwarz (BIC = - 2 logL + p log (N-r(X))), em que p é o
número de parâmetros do modelo; N é o total de observações e r é o posto
da matriz X (matriz de incidência para os efeitos fixos). O melhor ajuste
do modelo está em função dos menores valores desses critérios. Os AIC e
BIC permitem a comparação de modelos não hierárquicos e penalizam modelos
mais parametrizados, sendo o BIC mais rigoroso, favorecendo os modelos
mais parcimoniosos(14). Valores menores de AIC e BIC indicam
melhor ajuste do modelo.
Resultados e Discussão
A menor quantidade de registros para a característica peso
foi observada para o período do nascimento (dia um) até os 45 dias (Tabela
1). Por outro lado, foi verificado um aumento na quantidade de informações
em idades subsequentes, parte devido ao aumento na amplitude das classes
de idades utilizadas. O peso médio dos indivíduos teve um crescimento
linear de acordo com a idade. O maior coeficiente de variação foi
encontrado para a classe de 2 a 45 dias de idade, permanecendo constantes
nas demais classes utilizadas.
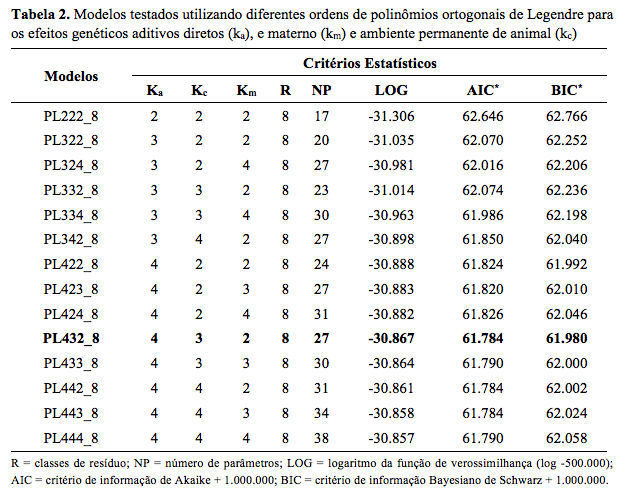
No presente trabalho foram testados 14 modelos com
diferentes ordens de ajuste dos polinômios ortogonais de Legendre para os
efeitos considerados (Tabela 2). O modelo de melhor ajuste, que apresentou
os menores valores para os critérios AIC e BIC, com um total de 27
parâmetros foi o PL432_8. Neste modelo, as variâncias genéticas aditivas
direta e materna e de ambiente permanente do animal foram modeladas com
polinômios de quarta, terceira e segunda ordem, respectivamente.
As estimativas das variâncias genéticas aditivas diretas
aumentaram gradualmente até os 500 dias de idade, sendo que, após esse
período, houve aumento acentuado nas estimativas (Figura 1a). Para o
ambiente permanente do animal, as variâncias estimadas no presente
trabalho aumentaram constantemente até próximo aos 400 dias de idade com
ligeira diminuição nas estimativas em idades posteriores (Figura 1b). A
estimativa de variância genética aditiva materna aumentou gradualmente de
forma linear do nascimento até o final do período estudado, porém menos
acentuada quando comparada com a variância genética aditiva direta (Figura
1a). Os resultados encontrados são semelhantes aos descritos por
Albuquerque e Meyer(15) para raça Nelore e por Sousa Júnior et
al.(16) e Dias et al.(17) para a raça Tabapuã;
entretanto, esses autores observaram aumento da variância genética aditiva
materna após a desmama dos animais. Os baixos valores de variância
genética aditiva materna observados no presente trabalho podem estar
associados à estrutura dos dados, na qual aproximadamente 80% das mães
possuíam apenas uma progênie. Segundo Ferreira(18), isso é
possível, pois estimativas de variância genética aditiva materna são
dependentes da proporção de elos mãe-progênie. Assim, quanto maior for
essa relação, maiores e mais acuradas são as estimavas de variância
genética aditiva materna.

A variância residual aumentou com a idade dos animais,
porém, os menores valores foram observados até os 180 dias de idade
(Figura 1d). É provável que a menor variância residual estimada no período
pré-desmama ocorra devido à homogeneidade de ambiente de criação dos
bezerros, fase em que o animal é basicamente dependente do ambiente
materno que, embora seja genético para a mãe, é um fator ambiental para o
bezerro. Contudo, deve-se considerar a variância em termos de proporção,
visto que a ordem de grandeza da variável é dependente da idade do animal.
Segundo Dias et al.(17), a ordem de ajuste dos polinômios e a
modelagem do resíduo podem interferir na estimação dos componentes de
variância. Dessa forma, as estimativas de parâmetros genéticos podem ser
influenciadas pelo modelo de regressão adotado e, portanto, a separação
dos efeitos aleatórios genéticos e de ambiente permanente em direto e
materno deve ser procedida para melhor modelar as mudanças que ocorrem no
crescimento de bovinos.
Quando a variância residual é homogênea, assume-se que o
ambiente temporário afetou igualmente todas as idades ao longo do
crescimento do animal, o que nem sempre ocorre. Além disso, pode ocorrer
um aumento da variância de ambiente permanente do animal. Por este motivo,
no presente estudo, as variâncias residuais foram modeladas
considerando-se oito classes diferentes como relatado por Huisman et al.(14).
Os autores reportaram que o mais adequado para ajustar pesos de bovinos ao
longo das idades é adotar estruturas heterogêneas de variância residual.
A herdabilidade direta estimada utilizando o modelo de
melhor ajuste (PL432_8) reduziu do nascimento até os 45 dias de idade,
aumentando após este período (Figura 2, à esquerda). O modelo de regressão
aleatória possibilitou estimar as herdabilidades em qualquer idade.

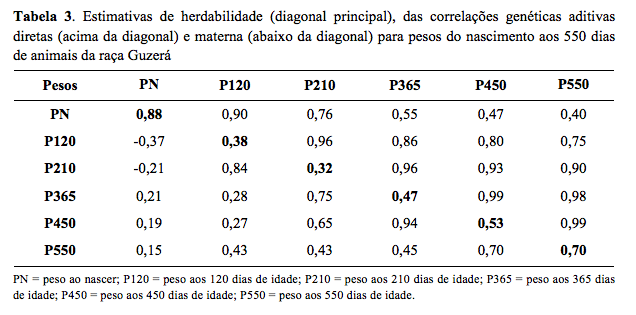
Considerando as idades-padrão (Tabela 3), as maiores
estimativas de herdabilidade direta foram observadas ao nascimento (0,88)
e aos 550 dias de idade (0,70), possivelmente devido ao menor número de
animais com informações nessas idades, o que pode ter comprometido as
estimativas dos componentes de variância. Entretanto, as estimativas para
os pesos aos 120 (P120), 210 (P210), 365 (P365) e 450 (P450) dias de idade
também foram expressivas, com tendência de aumento (0,38, 0,32, 0,47 e
0,53, respectivamente). Comportamento similar foi relatado por Sousa
Júnior et al.(16) para a raça Tabapuã, por Meyer(19)
para bovinos da raça Hereford e por Albuquerque e El Faro(20)
para a raça Nelore. Esses autores reportaram redução nas estimativas de
herdabilidade direta após o nascimento, aumentando com o avanço da idade
do animal. Resultados médios obtidos para a herdabilidade direta descritos
na literatura para P120, peso aos 240 dias (P240), peso ao ano (P365) e
P550 (0,23; 0,21; 0,30 e 0,41, respectivamente) foram menores aos
estimados no presente trabalho(21-23). Os valores das
herdabilidade direta sugerem que, na população estudada, existe
variabilidade genética suficiente para se obter ganho genético
considerável, por meio da seleção, principalmente a partir da desmama,
visto que as estimativas praticamente se mantiveram constantes e altas.
A herdabilidade materna estimada pelo melhor modelo
(PL432_8) aumentou do nascimento até os 45 dias de idade (0,10),
diminuindo gradativamente até os 550 dias de idade. A influência do efeito
materno na fase inicial de vida no animal foi expressiva até próximo aos
90 dias de idade, indicando que os maiores ganhos na seleção para
habilidade materna podem ser esperados próximos a essa idade. As
estimativas de herdabilidades maternas para peso ao nascimento (PN), P240,
P365 e P550 foram de 0,03; 0,05; 0,04 e 0,02, respectivamente. Os valores
médios de herdabilidade materna relatados por Ferraz Filho et al.(23),
Siqueira et al.(24) e Marcondes et al.(21) foram
superiores para P240 (0,19) e P365 (0,18), porém semelhantes para o P550
(0,02).
A variância de ambiente permanente de animal como proporção
da variância fenotípica diminuiu do nascimento, chegando à zero aos 15
dias de idade, seguido por um aumento até os 180 dias de idade (0,43),
próximo à fase de desmama dos animais (Figura 2, à direita). Após os 180
dias de idade, os valores foram decrescendo até os 550 dias de idade
(0,15). Comportamento semelhante foi relatado por Sousa Júnior et al.(16),
para a raça Tabapuã, utilizando modelos de regressão aleatória. As
estimativas de correlações genéticas aditivas diretas (Tabela 3) entre o
PN e pesos subsequentes foram positivas de alta magnitude, sendo maiores
em idades mais jovens, sugerindo que a seleção de animais para maiores
pesos em idades jovens ocasionará aumento do PN e, consequentemente, a
probabilidade de ocorrência de partos distócicos. Valores de correlações
genéticas maiores foram relatados por Boligon et al.(22) para a
raça Nelore entre PN e P240 (0,80) e o P550 (0,74), enquanto Albuquerque e
El Faro(20) encontraram valores menores entre PN e o peso aos
120 (0,77) e 205 dias de idade (0,45) também para a raça Nelore.
Esses valores de correlação genética sugerem que os genes
responsáveis pela expressão do peso em determinada idade são os mesmos
para as outras idades. Ou seja, a seleção para aumentar o peso pode ser
realizada em qualquer idade. No entanto, deve-se preconizar que a seleção
em idades jovens privilegia animais mais precoces e possivelmente a
maximização dos lucros dos sistemas de criação de bovinos de corte. Com
exceção das estimativas de correlações genéticas aditivas maternas de PN
com P120 (-0,37) e P210 (-0,21), todas as demais foram positivas, com
tendência de diminuição com o aumento da distância entre as idades.
Portanto, exceto para PN, quanto mais próximas as idades, maior a
correlação entre os efeitos maternos. Sousa Junior et al.(16) e
Albuquerque e El Faro(20) relataram comportamento similar para
bovinos da raça Nelore, pois observaram diminuição nos valores das
correlações genéticas entre o PN e pesos posteriores. A análise dos quatro
primeiros autovalores da matriz de covariância do efeito genético aditivo
direto explicaram 100% da variação genética aditiva total, sendo que cada
um foi responsável por 79,28%; 12,67%; 7,02% e 1,04% dessa variação,
respectivamente (Figura 3).
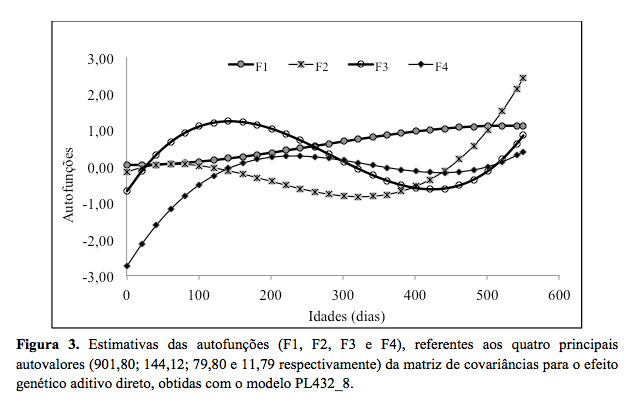
O primeiro autovalor explicou a maior parte da variação e
sua autofunção (F1) foi positiva para todas as idades. No entanto, segundo
Van der Werf(25), o sinal dos valores das autofunções não é
importante para a interpretação dos resultados; o que se deve considerar,
contudo, é a trajetória que as mesmas podem assumir durante o crescimento
do animal, pois indicam a magnitude do autovalor associado a cada
autofunção e as prováveis direções de seleção. Dessa forma, a seleção para
a primeira autofunção pode resultar em mudanças no mesmo sentido para a
característica peso em diferentes idades assim como observada nas
correlações genéticas. Comportamento semelhante foi observado por Boligon
et al.(26) em bovinos da raça Nelore, em que a seleção para a
primeira autofunção pode resultar em mudanças no mesmo sentido para os
pesos em diferentes idade. A segunda autofunção (F2) foi negativa até os
20 dias de idade, passando a ser positiva até os 100 dias de idade,
tornando-se negativa até os 440 dias de idade e posteriormente foi
positiva até os 550 dias de idade. A alteração no sinal sugere que a
seleção sobre este componente promoverá efeitos contrários no início e no
final da trajetória. Entretanto, considerando-se que o segundo autovalor
explica uma menor fração da variação genética aditiva (12,67%), a
possibilidade de mudança genética por seleção sobre esse componente pode
ser menor. Esses resultados apresentam o mesmo comportamento que os
descritos por Albuquerque e Meyer(15) em bovinos da raça
Nelore. A terceira e a quarta autofunção (F3 e F4) tiveram comportamento
oscilatório de negativo a positivo, sendo de pouca importância, uma vez
que juntas explicam pouca variância genética aditiva direta (8,06%) e,
portanto, as possibilidades de alteração genética por seleção com base em
suas autofunções pode ser limitada.
Conclusões
Modelos de regressão aleatória foram eficientes para
estimar, com grande acurácia, (co)variâncias e parâmetros genéticos, em
dados longitudinais de pesos de bovinos da raça Guzerá. Existe
variabilidade genética aditiva suficiente para promover ganhos genéticos
satisfatórios para o desempenho ponderal desde idades iniciais.
Agradecimentos
Os autores agradecem à Universidade Federal do Tocantins
pelo apoio financeiro (Edital Propesq/UFT nº 21/2014) e à ANCP pela
concessão dos dados.
Referências
1. Bertipaglia TS, Carreño LOD, Machado CHC, Andrighetto C,
Fonseca Rd. Estimates of genetic parameters for visual scores and their
correlation with production and reproductive traits in Brahman cattle.
Revista Brasileira de Zootecnia. 2012;41(6):1407-11.
2. Boligon A, Mercadante M, Baldi F, Lôbo R, Albuquerque L. Multi-trait
and random regression mature weight heritability and breeding value
estimates in Nelore cattle. South African Journal of Animal Science.
2009;39(5):145-8.
3. Kirkpatrick M, Hill WG, Thompson R. Estimating the covariance structure
of traits during growth and ageing, illustrated with lactation in dairy
cattle. Genetical research. 1994;64(01):57-69.
4. Meyer BK. Estimates of genetic and phenotypic covariance functions for
postweaning growth and mature weight of beef cows. Journal of Animal
Breeding and Genetics. 1999;116(3):181-205.
5. ALBUQUERQUE LG. Regressão aleatória: nova tecnologia pode melhorar a
qualidade das avaliações genéticas. 2004. In: Simpósio Brasileiro de
Melhoramento Animal, 5, 2004. Pirassununga, SP. Anais… Pirassununga: SBMA,
2004. (CD-ROM). Disponível em: http://sbmaonline.org.br/anais/v/palestras/pdfs/palest11.pdf,
Acesso em janeiro de 2017.
6. Van der Werf J, Schaeffer L. Random regression in animal breeding.
Course notes CGIL Guelph. 1997.
7. Albuquerque LGd, Meyer K. Estimates of covariance functions for growth
of Nelore cattle applying a parametric correlation structure to model
within-animal correlations. Livestock Production Science.
2005;93(3):213-22.
8. Meyer K. Random regressions to model phenotypic variation in monthly
weights of Australian beef cows. Livestock Production Science.
2000;65(1):19-38.
9. El Faro L, Albuquerque LGd. Utilização de modelos de regressão
aleatória para produção de leite no dia do controle, com diferentes
estruturas de variâncias residuais. Revista Brasileira de Zootecnia.
2003;32(5):1104-13.
10. Sakaguti E, Silva M, Martins E, Lopes P, Silva L, Quaas R, et al.
Trajetória de crescimento e efeito da idade da vaca nos modelos de
regressäo aleatória de bovinos jovens da raça Tabapuä. Arq bras med vet
zootec. 2002;54(4):414-23.
11. Helwig JT, Council K. Statistical analysis system user's guide. Carey,
NC, SAS Institute Inc. 1979:221-36.
12. Roso V, Schenkel F, editors. AMC-a computer programme to assess the
degree of connectedness among contemporary groups. Proceedings of the 8th
World Congress on Genetics Applied to Livestock Production, Belo
Horizonte, Minas Gerais, Brazil, 13-18 August, 2006; 2006: Instituto
Prociência.
13. Meyer K. WOMBAT—A tool for mixed model analyses in quantitative
genetics by restricted maximum likelihood (REML). Journal of Zhejiang
University Science B. 2007;8(11):815-21.
14. Huisman A, Veerkamp R, Van Arendonk J. Genetic parameters for various
random regression models to describe the weight data of pigs. Journal of
Animal Science. 2002;80(3):575-82.
15. Albuquerque LGd, Meyer K. Estimates of covariance functions for growth
from birth to 630 days of age in Nelore cattle. Journal of animal science.
2001;79(11):2776-89.
16. Sousa Júnior SCd, Oliveira SMPd, Albuquerque LGd, Boligon AA, Martins
Filho R. Estimação de funções de covariância para características de
crescimento da raça Tabapuã utilizando modelos de regressão aleatória.
Revista Brasileira de Zootecnia. 2010:1037-45.
17. Dias LT, Albuquerque LGd, Tonhati H, Teixeira RdA. Estimação de
parâmetros genéticos para peso do nascimento aos 550 dias de idade para
animais da raça Tabapuã utilizando-se modelos de regressão aleatória.
Revista Brasileira de Zootecnia. 2006;35(5):1915-25.
18. Ferreira JL. Efeito da estrutura dos dados sobre as estimativas de
(co) variâncias de peso à desmama em bovinos de corte, usando dados
simulados: Universidade Federal de Goiás; 2009 (Tese de doutorado).
Disponível em: https://repositorio.bc.ufg.br/tede/handle/tde/1177
19. Meyer K. Estimates of direct and maternal covariance functions for
growth of Australian beef calves from birth to weaning. Genetics Selection
Evolution. 2001;33(5):1.
20. de Albuquerque LG, ElFaro L. Comparações entre os valores genéticos
para características de crescimento de bovinos da raça Nelore preditos com
modelos de dimensão finita ou infinita. R Bras Zootec. 2008;37(2).
21. Marcondes C, Gavio D, Bittencourt T, Rocha J, Lôbo R, Bezerra L, et
al. Estudo de modelo alternativo para estimaçäo de componentes de (co)
variância e prediçäo de valores genéticos de características de
crescimento em bovinos da raça Nelore. Arq bras med vet zootec.
2002;54(1):93-9.
22. Boligon AA, ALBUQUERQUE Ld, Rorato PRN. Associações genéticas entre
pesos e características reprodutivas em rebanhos da raça Nelore. Revista
Brasileira de Zootecnia. 2008;37(4):596-601.
23. Ferraz Filho PB, de Amorim Ramos A, da Silva LOC, de Souza JC, de
Alencar MM, Malhado CHM. Tendência genética dos efeitos direto e materno
sobre os pesos à desmama e pós-desmama de bovinos da raça Tabapuã no
Brasil. R Bras Zootec. 2002;31(2):635-40.
24. de Siqueira RLPG, de Oliveira JA, Lôbo RB, Bezerra LAF, Tonhati H.
Análise da variabilidade genética aditiva de características de
crescimento na raça Nelore. R Bras Zootec. 2003;32(1):99-105.
25. Van der Werf J. 2001. Random regression in Animal Breeding: course
notes. Disponível em: http:
www-personal.une.edu.au=~jvanderw/CFcoursenotes.pdf. Acesso em: 20
dez. 2013.
26. Boligon AA, Mercadante MEZ, Forni S, Lôbo RB, Albuquerque LGd.
Covariance functions for weights from birth to maturity in Nellore cows.
Journal of Animal Science, 2010; 88:849-859./http: