
Resumo
Objetivou-se avaliar os efeitos de diferentes estratégias de alimentação restrita sobre as concentrações sanguíneas de hormônios, metabólitos e minerais de machos Holandês × Zebu castrados em terminação. Vinte machos Holandês × Zebu castrados (peso corporal inicial [PC] = 319 ± 16,2 kg; idade = 19 ± 1 meses) foram aleatoriamente distribuídos em 4 grupos de 5 animais, os quais receberam alimentação restrita a 85% do consumo ad libitum por 0 (R0; n=5), 28 (R28; n=5), 42 (R42; n=5) ou 84 (R84; n=5) dias. Os animais foram alojados individualmente e considerados como unidades experimentais. Amostras de sangue foram coletadas nos dias 0, 21, 42 e 84, e analizadas quanto às concentrações de fator de crescimento semelhante à insulina 1 (IGF-1), hormônios tireiodianos (T3 e T4), albumina, creatinina, ureia, proteína total, glicose, triglicerídeos, cálcio e fósforo. Animais que receberam alimentação restrita apresentaram aumento nas concentrações de IGF-1 até o dia 42 (P < 0,05). Animais R84 apresentaram as maiores (P < 0,05) concentrações de IGF-1 no dia 84. As concentrações de ureia foram maiores (P < 0,05) para os animais AL84 e menores (P < 0,05) para os animais R28; os animais R42 e R84 apresentaram valores intermediários e não diferentes (P > 0,05) entre si. As concentrações de IGF-1, ureia, albumina, e calcium, mas não as de T3, T4, glicose, triglicerídeos, creatinina, proteína total e fósforo, foram influencias pelas diferentes estratégias de alimentação restrita utilizadas em machos Holandês × Zebu castrados em terminação.
Palavras-chave: Balanço Mineral; Bovinos Cruzados; Consumo Alimentar Restrito
Abstract
The objectives were to determine the effects of restricted-feeding strategies on blood concentrations of hormones, metabolites, and minerals in Holstein × Zebu finishing steers. Twenty steers (body weight = 319 ± 16.2 kg; age = 19 ± 1 months) were randomly allotted in 4 groups of 5 steers, which were restricted to 85% ad libitum feed intake for either 0 (R0; n = 5), 28 (R28; n = 5), 42 (R42; n = 5) or 84 (R84; n = 5) days of an 84-d finishing period. Steers were individually housed and considered the experimental unit. Blood samples were collected on d 0, 21, 42, 63, and 84, and were assayed for insulin-like growth factor 1 (IGF-1), thyroid hormones (T3 and T4), albumin, creatinine, urea, total protein, glucose, triglycerides, calcium, and phosphorus. Steers had increasing IGF-1 concentrations until d 42 when restricted-fed. Steers fed R84 had the greatest IGF-1 concentrations on d 84. Urea concentrations were greatest in steers fed R0 and least in steers fed R28; steers fed R42 and R84 were intermediate and not different from one another. Concentrations of IGF-1, urea, albumin, and calcium, but not T3, T4, glucose, triglycerides, creatinine, total protein, and phosphorus were affected by restricted-feeding strategies in Holstein × Zebu finishing steers.
Keywords: Crossbred Cattle; Mineral Balance; Restricted Intake
Seção: Zootecnia
Recebido
6 de março de 2020.
Aceito
12 de agosto de 2020.
Publicado
19 de outubro de 202.
www.revistas.ufg.br/vet
Como citar - disponível no site, na página do artigo.
Introdução
Por definição, fornecer alimentação restrita é oferecer alimento em quantidades inferiores ao conumo ad libitum estimado(1). Na maioria dos casos, bovinos que recebem alimentação restrita durante a fase de crescimento apresentam crescimento compensatório (CC) quanto subsequentemente alimentados ad libitum na fase de terminação(2-4). Acredita-se que o CC ocorra devido à adaptação fisiológica prévia à restrição alimentar(5). No entanto, a ocorrência, a magnitude e a persistência das adaptações fisiológicas após o retorno à alimentação ad libitum são variáveis e dependem de fatores como raça, gênero, dieta, estágio de maturidade e severidade e duração da restrição alimentar(5-7). Assim, os mecanismos fisiológicos exatos do CC ainda não são totalmente compreendidos.
Os mecanismos fisiológicos em bovinos submetidos ao CC foram avaliados(8-10). Contudo, restrições alimentares severas (de 43 a 55 % do consumo ad libitum de MS) e/ou longos períodos (de 95 a 188 d) de restrição alimentar foram utilizados nesses estudos. Na maioria dos estudos, os bovinos que receberam alimentação restrita durante a fase de crescimento apresentaram desempenho produtivo e características de carcaça prejudicados ou necessitaram de tempo adicional em alimentação para atingir peso de abate quando comparados àqueles que foram continuamente alimentados ad libitum(3, 4, 11). Dessa forma, os dados sugerem que a alimentação restrita pode não ser aconselhável em condições práticas.
Em contraste, observou-se em um estudo anterior que a utilização de uma restrição alimentar moderada (15% do consumo ad libitum de MS) reduziu o consumo de alimentos, sem contudo prejudicar o desempenho produtivo ou as características de carcaça de machos Holandês × Zebu castrados em terminação, mesmo quando os animais receberam alimentação restrita durante toda a fase de terminação (84 d)(12). Nesse contexto, hipotetiza-se que o aumento da duração da restrição alimentar de 85% do consumo ad libitum de MS resultaria em maior magnitude e persistência das respostas fisiológicas experimentadas por machos Holandês × Zebu castrados em terminação. Assim sendo, objetivou-se avaliar os efeitos de diferentes estratégias de restrição alimentar sobre as concentrações sanguíneas de hormônios, metabólitos e minerais associados com o metabolismo do crescimento em machos Holandês × Zebu castrados em terminação ao longo do tempo.
Material e Métodos
Todos os procedimentos envolvendo animais seguiram os protocolos aprovados pelas normas da Comissão de Ética no Uso de Animais de Produção sob o protocolo 12/2016.
Foram utilizados vinte machos Holandês × Zebu castrados (idade média = 19 ± 1 meses; peso corporal inicial médio [PC] = 319 ± 16,2 kg). Os animais eram contemporâneos e oriundos de uma mesma fazenda. Antes do experimento, todos os animais foram criados em sistema de pastagem com suplementação. O experimento foi realizado utilizando o delineamento inteiramente casualizado. A duração total do experimento foi de 112 d, sendo 28 d para os animais se adaptarem ao local e às dietas e 84 d para coleta de dados. Os animais foram alojados em baias individuais cobertas de 6 m² com piso de concreto, equipadas com comedouros individuais e bebedouros de concreto de fluxo contínuo. A temperatura média do ar, umidade e precipitação durante o ensaio foram de 18ºC, 79% e 7 mm, respectivamente.
No final do período de adaptação, os animais foram pesados após jejum de sólidos de 14 horas e aleatoriamente distribuídos em 1 de 4 tratamentos (n = 5 animais por tratamento): alimentação ad libitum por 84 d (R0); alimentação restrita durante os primeiros 28 d (R28); alimentação restrita durante os primeiros 42 d (R42); ou alimentação restrita durante 84 d (R84). Os animais R28, R42 e R84 receberam alimentação restrita a 85% do consumo ad libitum de matéria seca, com base no consumo de matéria seca (CMS) dos animais R0, durante os respectivos períodos de restrição alimentar. Animais R28 e R42 tiveram acesso a alimentação ad libitum pelo restante do período de terminação, por 56 e 42 d, respectivamente.
A dieta experimental foi a mesma independentemente do tratamento. A dieta foi calculada de acordo com as recomendações do BR CORTE(13) a fim de fornecer aproximadamente 11% de proteína bruta (PB) com base na MS. A dieta era composta por 40% de silagem de milho e 60% de concentrado (com base na MS). O concentrado era composto por milho grão moído, farelo de soja, ureia, sulfato de amônio, bicarbonato de sódio, óxido de magnésio, sal comum, calcário e mistura mineral (Tabela 1).
A silagem de milho e o concentrado foram pesados separadamente e depois misturados à mão no momento da alimentação. A dieta completa foi fornecida uma vez ao dia (0800 h) e os animais tiveram livre acesso a água limpa. A dieta fornecida para os animais alimentados ad libitum foi ajustada diariamente para que as sobras fossem mantidas em aproximadamente 2% do total oferecido com base na matéria natural, sendo as sobras misturadas com a nova dieta oferecida no dia seguinte. Os animais foram pesados a cada 14 d para monitorar o ganho médio diário (GMD) e ajuste do CMS animais recebendo alimentação restrita. Maiores informações sobre os procedimentos experimentais do estudo e sobre o efeito dos tratamentos sobre CMS, PC, GMD, eficiência alimentar, características de carcaça e crescimento de componentes não-carcaça foram previamente descritos por Silva et al.(12).
Foram coletadas três amostras de sangue de cada um dos animais nos d 0, 21, 42, 63 e 84. As amostras de sangue foram coletadas por punção da veia jugular às 0700 h, antes da alimentação. Duas das amostras foram coletadas em tubos de 8,5 mL com ativador de coágulo e gel para separação de soro (BD Vacuntainer® SST II Plus, São Paulo, Brasil) para análise de triiodotironina total (T3), tiroxina total (T4), crescimento semelhante à insulina fator 1 (IGF-1), albumina, ureia, proteína total, creatinina, triglicerídeos, fósforo (P) e cálcio (Ca). A terceira amostra foi coletada em tubo de 6 mL com EDTA e fluoreto de sódio (BD Vacutainer® Fluoreto/EDTA, São Paulo, Brasil) para análise da glicose. Após a coleta, as amostras foram centrifugadas a 3.600 × g por 15 min a 4 ° C, em seguida o soro e plasma foram transferidos para microtubos de 2 mL e imediatamente congelados a -80°C para posteriores análises.
As concentrações séricas totais de T3 e T4 foram determinadas nos d 21, 42, 63 e 84. As concentrações séricas totais de T3 e T4 foram analisadas por imunoensaio de quimioluminescência utilizando-se kits da Beckman Coulter (ref. 33830 e 33800, respectivamente; Beckman Coulter®, Brea, EUA), no Access 2 Immunoassay System (Beckman Coulter Inc., Brea, EUA). O IGF-1 foi quantificado usando os kits DiaSorin (ref. 313231; DiaSorin®, Saluggia, Itália) em analisador automatizado de quimioluminescência (Liaison®, Saluggia, Itália).
As concentrações plasmáticas de glicose foram determinadas utilizando um kit (ref. K082) da Bioclin Diagnostics (Bioclin, Quibasa Química Básica Ltda, Belo Horizonte, Brasil). As concentrações séricas de triglicerídeos (ref. K117), uréia (ref. K056), proteína total (ref. K031); creatinina (ref. K067) e albumina (ref. K040) foram determinadas utilizando-se kits da Bioclin Diagnostics (Bioclin® Quibasa Química Básica Ltda, Belo Horizonte, Brasil). As concentrações séricas de Ca (ref. K051) e P (ref. K068) foram quantificadas utilizando-se kits da Bioclin Diagnostics (Bioclin® Quibasa Química Básica Ltda, Belo Horizonte, Brasil). Os metabólitos e minerais descritos anteriormente foram analisados de acordo com instruções do fabricante em um analisador bioquímico automático (Mindray BS200E, Shenzhen Mindray Bio-Medical Electronics Co. Ltd., Shenzhen, China).
Os dados referentes às concentrações sanguíneas de hormônios, metabólitos e minerais foram analisados em delineamento inteiramente casualizado, utilizando-se o procedimento PROC MIXED no SAS (versão 9.4, SAS Institute Inc., Cary, NC, EUA). Tratamento e dia de amostragem, bem como sua interação, foram considerados efeitos fixos no modelo. Medidas repetidas foram utilizadas para analisar o dia de amostragem. A estrutura de covariância foi escolhida com base no menor valor para o critério de informação de Akaike corrigido (AICC) para modelos executados sob simetria composta, simetria composta heterogênea, autorregressiva de primeira ordem, matriz em bandas ou estruturas de variância-covariância não estruturadas. Os graus de liberdade foram estimados pelo método Kenward-Roger. Os animais foram as unidades experimentais. As concentrações sanguíneas do hormônio, metabólito ou mineral no d 0 foram utilizadas como covariáveis, exceto para as concentrações totais de T3 e T4, uréia e Ca, porque estas não foram significativas para as concentrações de uréia e Ca, e nenhum dado estava disponível no d 0 para as concentrações totais de T3 e T4. As médias foram comparadas utiliza-se o teste Diferença Mínima Significativa de Fisher (DMS). Declarou-se significância quando P ≤ 0,05, e tendência quando 0,05 < P ≤ 0,10.
Resultados
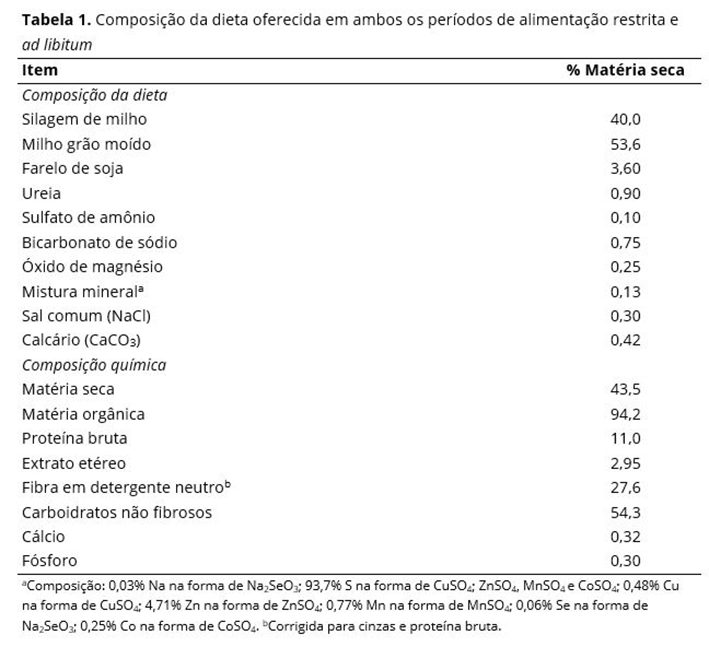
Observou-se interações tratamento × dia (P ≤ 0,05) para as concentrações de IGF-1 (Figura 1a) e Ca (Figura 1b). Os valores de IGF-1 (Figura 1a) foram diferentes entre os dias de amostragem (P < 0,02) para todos os tratamentos, exceto para animais R0 (P ≥ 0,36). Os animais recebendo alimentação restrita apresentaram concentrações de IGF-1 crescentes até o dia 42 (P < 0,05). Enquanto as concentrações de IGF-1 foram mais variáveis nos d 63 e 84. Contudo, diferenças (P < 0,05) entre os tratamentos dentro de cada dia de amostragem somente foram observadas no d 84, onde os níveis de IGF-1 foram maiores (P < 0,05) para animais R84 e menores para animais R0 e R28; animais R42 apresentaram valores intermediários e semelhantes (P > 0,05) aos demais tratamentos.
Não foram observadas diferenças (P ≥ 0,34) entre os tratamentos para as concentrações de Ca nos d 0, 21, e 42 (Figura 1b). No entanto, as concentrações de Ca foram superiores (P < 0,05) em animais R0, R28, e R42 quando comparados aos animais R84 no d 63. No d 84, maiores (P < 0,05) concentrações de Ca coforam observadas para animais R0 e R28 em relação aos animais R42 e R84.
Houve uma tendência de interação tratamento × dia (P ≤ 0,08) para as concentrações de P (Figura 1c). No d 63, os valores de P tenderam a serem maiores (P < 0,10) nos animais R42 e menores nos animais R28; animais R0 e R84 tenderam a serem intermediários e não diferentes (P > 0,10) dos animais dos demais tratamentos (Figure 1c). Além disso, no d 84, as concentrações de P tenderam a serem maiores (P < 0,10) nos animais R84 e menores nos animais R28; animais R0 e R42 tenderam a serem intermediários e não diferentes (P > 0,10) dos animais dos demais tratamentos.
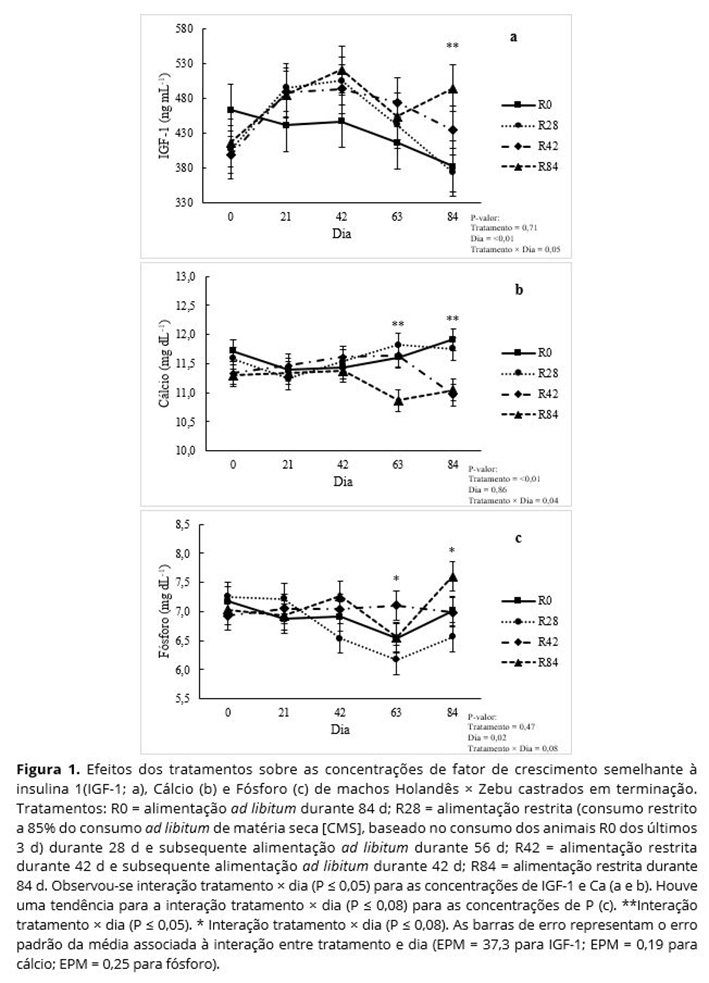
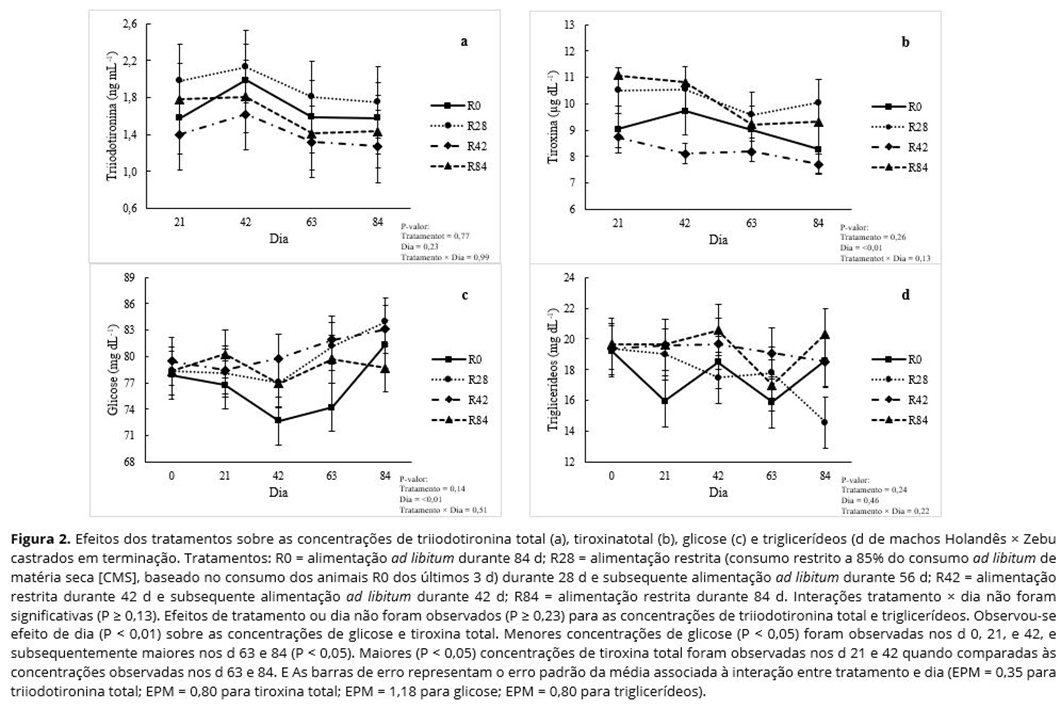
Não foram observadas interações tratamento × dia (P ≥ 0,13) para as concentrações de T3 (Figura 2a) e T4 (Figura 2b) totais, glicose (Figura 2c), triglicerídeos (Figura 2d), ureia (Figura 3a), creatinina (Figura 3b), albumina (Figura 3c) ou proteina total (Figura 3d). Não foram observados efeitos de tratamento ou dia (P ≥ 0,23) para as concentrações de T3 total (Figura 2a), creatinina (Figura 3b), triglicerídeos (Figura 2d) ou proteína total (Figura 3d).
As concentrações de ureia foram maiores (P < 0,05) em animais R0 e menores (P < 0,05) em animais R28; animais R42 e R84 foram intermediários e não diferentes (P > 0,05) entre si (Figura 3a). As concentrações de albumina foram maiores (P < 0,05) nos animais R42 e R84, e menores nos animais R0; animais R28 foram intermediários e não diferentes (P > 0,05) dos animais dos demais tratamentos (Figura 3c). Embora não tenha sido observado efeito de tratamento sobre as concentrações de glicose e T4 total (P ≥ 0,14), observou-se um efeito de dia (P ≤ 0,01) para os níveis de glicose (Figura 2c) e T4 total (Figura 2b). As concentrações de glicose não diferiram (P > 0,05) nos d 0, 21, e 42, mas aumentou no d 84 (P < 0,05), independentemente do tratamento. As concentrações de glicose no d 63 foram intermediárias e não diferentes dos demais dias (P > 0,05). Maiores (P < 0.05) concentrações de T4 total foram observadas nos d 21 e 42 quando comparadas às concentrações nos d 63 e 84, independentemente do tratamento.
Discussão
As concentrações sanguíneas de hormônios, metabólitos e minerais refletem o status nutricional do animal(8). Dessa forma, são esperadas alterações em suas concentrações durante períodos de restrição alimentar em ruminantes, como resposta à redução do consumo de alimentos, pois o animal está tentando manter a homeostase(5, 14). A alteração pode ser revertida dentro de alguns dias após o retorno à alimentação ad libitum, dependendo da severidade e duração da restrição alimentar(8, 10, 15).
Por exemplo, níveis reduzidos de glicose, hormônios tireoidianos e IGF-1 têm sido relatados em bovinos recebendo alimentação restrita(8, 10, 16), como resultado da redução do consumo de alimentos. Os pesquisadores teorizam que esta resposta seja uma tentativa de reduzir o metabolismo basal e o gasto de energia e alterar a utilização preferencial de substratos para manter a homeostase, ao invés de estimular o crescimento(17, 18). Maiores concentrações transitórias (no início do período de restrição alimentar) ou persistentes (durante todo o período de restrição alimentar) de ácidos graxos não esterificados, beta-hidroxibutirato, ureia, creatinina, albumina e proteína total também foram relatadas durante a restrição alimentar(9, 10, 15). Essas pesquisas sugeriram que houve uma mobilização de gordura e proteína para superar o déficit de energia quando os bovinos foram alimentados com restrição de 43 a 55% do CMS ad libitum de 95 a 180 d(9, 10, 15).
No entanto, os resultados apresentados no presente trabalho não seguiram este mesmo padrão. Não foram observadas interações tratamento × dia para qualquer um dos metabólitos avaliados. Além disso, nenhum efeito de tratamento foi observado para as concentrações de glicose, triglicerídeos, creatinina, proteína total ou T3 e T4 totais; e os níveis de IGF-1 não foram reduzidos durante o período de restrição alimentar nos animais que receberam alimentação restrita. Os resultados apresentados sugerem que a restrição alimentar de 15% CMS imposta no presente estudo não foi severa o suficiente para reduzir o metabolismo basal ou prejudicar o estímulo ao crescimento.
Os dados apresentados no presente estudo podem conflitar com trabalhos anteriores porque, apesar do CMS reduzido, animais continuaram a crescer, embora a uma taxa ligeiramente reduzida (1,40 vs. 1,30 kg d-1), durante os períodos de alimentação restrita(12). Nos estudos previamente descritos, quando os bovinos foram severamente restritos, o consumo de alimentos foi suficiente apenas para manutenção do PC (9, 16) ou permitindo crescimento mínimo (de 0,12 to 0,6 kg d-1)(8, 10, 15), enquanto os animais do presente estudo apresentaram GMD de aproximadamente 1,30 kg d-1 durante os períodos de alimentação restrita(12).
A duração da alimentação restrita afetou as concentrações de ureia e albumina. A concentração de ureia é um indicador do status proteico em bovinos, pois está positivamente associada com a ingestão de nitrogênio (N) e o turnover proteico(19, 20). Todos os animais do presente estudo foram alimentados com a mesma dieta; assim, o consumo de N diferiu entre os tratamentos, visto que o CMS foi atribuído de acordo com o tratamento. Dessa forma, as maiores concentrações de ureia observadas nos animais R0 quando comparados aos animais R28, R42 e R84 poderiam ser resultado do maior consumo de N dietético(12).
Por outro lado, os menores valores de ureia observados nos animais R28, R42 e R84 em relação ao animais R0 também poderiam ser atribuídos ao aumento da eficiência de uso de N quando os animais receberam alimentação restrita(5, 10, 15). Além disso, entre animais que receberam alimentação restrita, as menores concentrações de ureia foram observadas para os animais R28 quando comparados aos animais R42 e R84, sugerindo que a eficiência do uso de N foi maior quando os bovinos foram submetidos à restrição por um curto período de tempo. O fato de que o desempenho produtivo e as características de carcaça não foram prejudicados mesmo quando animais receberam alimentação restrita durante toda a fase de terminação (84 d); e que os animais R28 apresentaram a maior retenção corporal de N entre todos os tratamentos(12), sugere que houve um aumento da eficiência do uso de N nos animais que passaram por restrição alimentar.
O nível de albumina também é um indicador do status proteico em bovinos(21, 22). No presente trabalho, as concentrações de albumina aumentaram à medida que a duração da restrição alimentar aumentou, e as maiores concentrações foram observadas quando os animais receberam alimentação restrita por 42 e 84 d. Keogh et al.(10) observaram maiores concentrações de albumina nos estágios iniciais da restrição alimentar em machos Holandês não castrados. Os autores deste estudo presumiram que esse resultado refletia a utilização de reservas proteicas corporais para superar o déficit de energia dos animais. Contudo, como discutido anteriormente, a restrição alimentar imposta no presente estudo não foi severa o suficiente para causar um balanço energético negativo, independentemente da duração da restrição. Assim, no presente estudo, o aumento do catabolismo proteico para atender às necessidades energéticas dos animais não explica o aumento dos valores de albumina nos animais que receberam alimentação restrita quando comparados àqueles que receberam alimentação ad libitum. No entanto, os autores não têm conhecimento de nenhum estudo que tenha associado o aumento das concentrações de albumina com o aumento do anabolismo proteico.
O IGF-1 é bem caracterizado como promotor do crescimento muscular(17, 18). A concentração sérica de IGF-1 está diretamente relacionada com os status nutricional de ruminantes. Nesse contexto, sua concentração aumenta com o aumento do consumo de alimentos e diminui em condições de restrição alimentar(23, 24). A redução nos níveis circulantes de IGF-1 durante o período de restrição alimentar foi relatada anteriormente em fêmeas(9, 25), machos castrados(8, 15) e machos não castrados(3, 10). Este efeito da restrição alimentar foi revertido alguns dias após o retorno à alimentação ad libitum. No entanto, no presente estudo, 15% de restrição do CMS não promoveu redução das concentrações de IGF-1, independentemente da duração da restrição. Pelo contrário, as concentrações de IGF-1 aumentaram à medida que a duração da restrição aumentou, sugerindo que a produção de IGF-1 foi estimulada quando uma restrição alimentar moderada é utilizada. Quando o acesso à alimentação ad libitum foi garantido, as concentrações de IGF-1 dos animais R28 gradualmente retornaram aos níveis observados no d 0. Contudo, no d 84, as concentrações de IGF-1 permaneceram maiores em animais R42 quando comparados aos valores observados no d 0 e para os animais R0, e animais que receberam alimentação restrita durante todos os 84 d (R84) apresentaram as maiores concentrações de IGF-1.
O estímulo das células musculares pelo IGF-1 resulta no aumento da captação de aminoácidos e glicose, proliferação das células satélites e síntese proteica, além da diminuição da proteólise e apoptose(17, 18, 26). Assim, as menores concentrações de ureia em combinação com as maiores concentrações de IGF-1 nos animais que receberam alimentação restrita indicam que o IGF-1 poderia estar melhorando a conservação de N e o crescimento muscular.
Diferentemente da maioria dos estudos anteriores, os efeitos da duração da alimentação restrita sobre as concentrações de Ca e P também foram avaliados no presente trabalho. As concentrações de Ca e P são estreitamente reguladas através de mecanismos homeostáticos(27, 28). Assim, exceto em casos extremos, as concentrações séricas de Ca e P são mantidas relativamente constantes, entre 9 e 11 e 6 e 8 mg dL-1, respectivamente(29, 30), o que torna difícil interpretar os efeitos de tratamentos dietéticos em suas concentrações sanguíneas.
A dieta utilizada no presente estudo supriu aproximadamente 32 e 26 g d-1 de Ca quando os animais receberam alimentação ad libitum e restrita, respectivamente. A dieta supriu 30 e 25 g d-1 P quando os animais receberam alimentação ad libitum e restrita, respectivamente. De acordo com o BR-CORTE(13), as exigências diárias de machos Holandês × Zebu castrados em terminação com PC médio de 319 kg e GMD de 1,38 kg d-1 (média dos animais do presente estudo) são 32,13 e 19,29 g d-1 de Ca e P, respectivamente. Dessa forma, apesar do menor CMS, o suprimento de P foi aproximadamente 5.7 g acima das exigências diárias; contudo, os animais estavam tendo um déficit diário de aproximadamente 6 g de Ca durante o período de restrição alimentar. Apesar do fornecimento insuficiente de Ca dietético quando os animais receberam alimentação restrita, os níveis de Ca permaneceram dentro da faixa normal para bovinos(22, 31) durante todo o experimento, 10,9 e 11,8 mg dL-1. Este resultado não é surpreendente, pois os mecanismos homeostáticos para manter as concentrações de Ca circulantes são estreitamente regulados(32). Essa estreita regulação provavelmente reduziu a variação nas concentrações de Ca e P, fazendo com que diferenças significativas fossem mais facilmente observadas quando pequenas variações numéricas dentro e entre os tratamentos ocorreram.
As concentrações de glicose e T4 total não diferiram entre os tratamentos. No entanto, observou-se efeito de dia de coleta para os valores de glicose e T4 total, onde animais de todos os tratamentos apresentaram concentrações de glicose aumentadas e T4 total reduzidas nos d 63 e 84. O aumento dos níveis de glicose próximo ao final do experimento pode ter sido resultado de uma redução da sensibilidade à insulina à medida que os animais ganharam peso e ficaram mais pesados(33, 34). Frequentemente, as concentrações de T4 são positivamente correlacionadas com a taxa metabólica, e sabe-se que a taxa metabólica diminui com o avanço da idade ou conforme os animais atingem a maturidade(35-37). Os sistemas de produção em confinamento de países em desenvolvimento são diferentes daqueles usados em países desenvolvidos. Nos Estados Unidos, por exemplo, os animais são confinados com idades entre 7 e 16 meses por 140 a 180 d, em média(38). No Brasil, no entanto, os bovinos são criados primeiramente em pastagens por 18 a 21 meses e depois são transferidos para confinamentos onde permanece por cerca de 80 a 100 d, sendo abatidos com idades de 21 a 25 meses(39). No presente estudo, animais tinham 19 meses de idade no início do período de confinamento, quando o crescimento do tecido ósseo está praticamente concluído e uma quantidade razoável de tecido muscular já foi depositada. Ou seja, os animais estavam mais próximos da maturidade do que os bovinos comumente terminados em confinamentos nos Estados Unidos, os quais são abatidos com idade entre 16 e 20 meses. Assim, a redução das concentrações de T4 próximo ao final do período de confinamento pode ter sido resultado do processo natural de envelhecimento e maturidade.
Conclusões
Este trabalho demonstrou os possíveis mecanismos pelos quais machos Holandês × Zebu castrados em terminação se adaptam a estratégias de restrição alimentar moderada. Independentemente da duração da restrição alimentar (15% de restrição do CMS), os resultados apresentados no presente estudo não indicaram mudanças na mobilização de tecidos para suprir o déficit de energia ou redução do metabolismo basal e gasto de energia em machos Holandês × Zebu castrados em terminação. Contudo, os resultados também sugeriram aumento da eficiência de utilização de N e do crescimento muscular quando os animais receberam alimentação restrita. Estes resultados podem contribuir para um melhor manejo alimentar de bovinos em confinamento.
Agradecimentos
Este estudo e bolsa do primeiro autor foram financiados pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ, Brasil), pela Coordenação de Pessoal de Nível Superior (CAPES, Brasil), pela Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, Brasil) e pelo Instituto Nacional de Ciência e Tecnologia em Ciência Animal (INCT – CA, Brasil). Agradecimentos também são estendidos ao Department of Animal Science of Pennsylvania State University pela colcaboração no preparo deste manuscrito.
Referências
1.Galyean ML. Review: Restricted and programmed feeding of beef cattle-definitions, application, and research results. PAS. 1999; 15:1–6.
2.Carstens GE, Johnson DE, Ellenberger MA, Tatum JD. Physical and chemical components of the empty body during compensatory growth in beef steers. J Anim Sci. 1991; 69:3251–64.
3.Hornick JL, Van Eenaeme C, Diez M, Minet V, Istasse L. Different periods of feed restriction before compensatory growth in Belgian Blue bulls: II. Plasma metabolites and hormones. J Anim Sci. 1998; 76:260–71.
4.Felix TL, Radunz AE, Loerch SC. Effects of limit feeding corn or dried distillers grains with solubles at 2 intakes during the growing phase on the performance of feedlot cattle. J Anim Sci. 2011; 89:2273–79.
5.Hornick JL, Van Eenaeme C, Gérard O, Dufrasne I, Istasse L. Mechanisms of reduced and compensatory growth. Domest Anim Endocrinol. 2000; 19:121–32.
6.Wilson PN, Osbourn DF. Compensatory growth after undernutrition in mammals and birds. Biol Rev. 1959; 35:324–61.
7.Mahyuddin, P. Compensatory growth in ruminants. Anim Prod. 2004; 6:125–35.
8.Ellenberger MA, Johnson DE, Carstens GE, Hossner KL, Holland MD, Nett TM, Nockels CF. Endocrine and metabolic changes during altered growth rates in beef cattle. J Anim Sci. 1989; 67:1446–54.
9.Yambayamba ES, Price MA, Foxcroft GR. Hormonal status, metabolic changes, and resting metabolic rate in beef heifers undergoing compensatory growth. J Anim Sci. 1996; 74:57–69.
10.Keogh K, Waters SM, Kelly AK, Wylie A RG, Sauerwein H, Sweeney T, Kenny DA. Feed restriction and realimentation in Holstein-Friesian bulls: II. Effect on blood pressure and systemic concentrations of metabolites and metabolic hormones. J Anim Sci. 2015; 93:3590–3601.
11.Keogh K, Waters SM, Kelly AK, Kenny DA. Feed restriction and subsequent realimentation in Holstein Friesian bulls: I. Effect on animal performance; muscle, fat, and linear body measurements; and slaughter characteristics. J Anim Sci. 2015; 93:3578–89.
12.Silva FAS, Valadares Filho SC, Godoi LA, Silva BC, Pacheco MVC, Zanetti D, Benedeti PDB, Silva FF, Felix TL. Effect of duration of restricted-feeding on nutrient excretion, animal performance, and carcass characteristics of Holstein × Zebu finishing steers. Anim Prod Sci. 2020; 60:535–544.
13.Valadares Filho SC, Costa e Silva LF, Gionbelli MP, Rotta PP, Marcondes MI, Chizzotti ML, Prados LF. Nutrient Requirements of Zebu and Crossbred Cattle - BR-CORTE. 3rd ed. Visconde do Rio Branco: Suprema Gráfica Ltda.; 2016. 314p. English.
14.Hoch T, Begon C, Picard B. Mécanismes et conséquences de la croissance compensatrice chez les ruminants. INRA Prod Anim. 2003; 16:49–59.
15.Hayden JM, Williams JE, Collier RJ. Plasma growth hormone, insulin-like growth factor, insulin, and thyroid hormone association with body protein and fat accretion in steers undergoing compensatory gain after dietary energy restriction. J Anim Sci. 1993; 71:3327-3338.
16.Blum J W, Schnyder W, Kunz PL, Blom AK, Bickel H, Schürch A. Reduced and compensatory growth: Endocrine and metabolic changes during food restriction and refeeding in steers. J Nutr. 1985; 115:417–24.
17.Breier BH. Regulation of protein and energy metabolism by the somatotropic axis. Domest Anim Endocrinol. 1999; 17:209–18.
18.Hossner KL. Hormonal regulation of farm animal growth. Cambridge: CABI Publishing; 2005. 240p. English.
19.Preston R, Schnakenberg D, Pfander W. Protein utilization in ruminants: I. Blood urea nitrogen as affected by protein intake. J Nutr. 1965; 86:281–88.
20.Kohn RA, Dinneen MM, Russek-Cohen E. Using blood urea nitrogen to predict nitrogen excretion and efficiency of nitrogen utilization in cattle, sheep, goats, horses, pigs, and rats. J Anim Sci. 2005; 83:879–89.
21.Russell KE, Roussel AJ. Evaluation of the ruminant serum chemistry profile. Vet Clin North Am Food Anim Pract. 2007; 23:403–26.
22.Kaneko JJ, Harvey JW, Bruss ML. Clinical biochemistry of domestic animals. 6th ed. San Diego: Academic Press; 2008. 928p. English.
23.Bass JJ, Oldham JM, Hodgkinson SC, Fowke PJ, Sauerwein H, Molan P, Breier BH, Gluckman PD. Influence of nutrition and bovine growth hormone (GH) on hepatic GH binding, insulin-like growth factor-I and growth of lambs. J Endocrinol. 1991; 128:181–86.
24.Mcguire MA, Vicini JL, Bauman DEP, Veenhuizent JJ. Insulin-like growth factors and binding proteins in ruminants and their nutritional regulation. J Anim Sci. 1992; 70:2901–10.
25.Ronge H, Blum J. Insulin-like growth factor I responses to recombinant bovine growth hormone during feed restriction in heifers. Acta Endocrinol. 1988; 120:735–44.
26.Oksbjerg N, Gondret F, Vestergaard M. Basic principles of muscle development and growth in meat-producing mammals as affected by the insulin-like growth factor (IGF) system. Domest Anim Endocrinol. 2004; 27:219–240.
27.Rowlands GJ. A review of variations in the concentration of metabolites in the blood of beef and dairy cattle associated with physiology, nutrition and disease, with particular reference to the interpretation of metabolic profiles. World Rev Nutr Diet. 1980; 35:172–235.
28.Reinhardt TA, Horst RL, Goff JP. Calcium, phosphorus, and magnesium homeostasis in ruminants. Vet Clin North Am Food Anim Pract. 1988; 4:331–50.
29.NRC. Nutrient requirements of beef cattle. 7th ed. Washington: Nat. Acad. Press; 2000. 248p. English.
30.Berchielli TT, Pires AV, Oliveira SG. Nutrição de Ruminantes. 2nd ed. Jaboticabal: Editora Funep; 2011. 616p. Portuguese.
31.Doornenbal H, Tong AKW, Murray NL. Reference values of blood parameters in beef cattle of different ages and stages of lactation. Can J Vet Res. 1988; 52:99–105.
32.McDowell LR. Minerals in Animal and Human Nutrition. 2nd ed. Amsterdam: Elsevier; 2003. 660p. English.
33.Eisemann JH, Huntington GB, Catherman DR. Insulin sensitivity and responsiveness of portal-drained viscera, liver, hindquarters, and whole body of beef steers weighing 275 or 490 kilograms. J Anim Sci. 1997; 75:2084–91.
34.Radunz AE, Fluharty FL, Relling AE, Felix TL, Shoup LM, Zerby HN, Loerch S C. Prepartum dietary energy source fed to beef cows: II. Effects on progeny postnatal growth, glucose tolerance, and carcass composition. J Anim Sci. 2012; 90:4962–74.
35.Kleiber M. Body size and metabolic rate. Physiol Rev. 1947; 27:511–41.
36.Rolfe DFS, Brown GC. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol Rev. 1997; 77:731–58.
37.Hulbert AJ, Pamplona R, Buffenstein R, Buttemer WA. Life and death: metabolic rate, membrane composition, and life span of animals. Physiol Rev. 2007; 87:1175–1213.
38.Samuelson KL, Hubbert ME, Galyean ML, Löest CA. Nutritional recommendations of feedlot consulting nutritionists: The 2015 New Mexico State and Texas Tech University survey. J Anim Sci. 2016; 94, 2648–63.
39.Ferraz JBS, Felício PE. Production systems - An example from Brazil. Meat Sci. 2010; 84:238–43.