
Resumo
A aplicação de biotecnologias na reprodução animal tem possibilitado produzir formas jovens em quantidade e qualidade. O aumento da quantidade de gametas viáveis produzidos pelos reprodutores, mediante uma dieta ideal, entre outros fatores, pode garantir uma maior produção. Assim, o objetivo deste trabalho foi determinar o efeito de três dietas quanto à sobrevivência espermática de Macrobrachium acanthurus. Para tanto, 24 machos de M. acanthurus foram utilizados, sendo distribuídos ao acaso e de forma igual entre os tratamentos. As dietas foram compostas por 100% de alimento fresco (músculo de peixe e lula - 14% de proteína), 100% de alimento seco (ração comercial - 50% de proteína) e uma mescla dessas dietas contendo 30% de proteína. Os espermatóforos foram extraídos por eletroestimulação a cada 15 dias, sendo o controle aqueles obtidos diretamente da natureza. Não houve diferença significativa entre as dietas quando comparado os pesos dos camarões e dos espermatóforos. A alimentação 100% fresca proporcionou o melhor desempenho para a sobrevivência espermática.
Palavras-chave: Caridae; camarão canela; nutrição; esperma
Abstract
The application of biotechnology in animal reproduction has enabled the production of young forms in both quantity and quality. Increasing the number of viable gametes produced by reproducers, among other factors, through an ideal diet, can ensure higher production. Therefore, the aim of this study was to determine the influence of three diets on the sperm survival of Macrobrachium acanthurus. To this end, 24 M. acanthurus males were used, distributed randomly and equally among treatments. Their diets were composed of 100% fresh food (fish and squid muscle - 14% protein), 100% dry feed (commercial feed - 50% protein) and a mixture of these diets containing 30% protein. Spermatophores were extracted through electrical stimulation every 15 days, and the controls consisted of spermatophores obtained directly from nature. No significant difference between diets was observed comparing shrimp and spermatophore weights. The 100% fresh diet provided the best sperm survival performance.
Keywords: Caridae; cinnamon shrimp; nutrition; sperm
Seção: Recursos Pesqueiros e Engenharia de Pesca
Recebido
20 de agosto de 2019
Aceito
28 de janeiro de 2020.
Publicado
9 de setembro de 2020.
www.revistas.ufg.br/vet
Como citar - disponível no site, na página do artigo.
Introdução
Para o crescimento do cultivo de camarão, foram necessários investimentos na produção, manejo, nutrição, reprodução e, mais recentemente, na melhoria genética das espécies. Manter camarões com baixa qualidade reprodutiva implica altos custos para os aquicultores. Algumas técnicas para melhorar a qualidade reprodutiva têm sido realizadas em Litopenaeus vannamei e M. rosenbergii, como a avaliação de características da gônoda ou moléculas relacionadas funcionalmente com o processo reprodutivo(5).
De acordo com Browdy(6), Meumpol et al.(7) e Shailender et al.(8), as dietas e a qualidade dos reprodutores interferem diretamente na produção dos gametas e estes são responsáveis pela boa qualidade das larvas, sendo importante para o êxito do cultivo. Em geral, os efeitos nutricionais sobre a reprodução de camarões têm sido concentrados principalmente em fêmeas. No entanto, os machos podem estar contribuindo para um êxito limitado de algumas espécies nativas visando à criação em cativeiro(9-13).
Em machos, a qualidade espermática pode ser uma ferramenta para avaliar a eficiência da dieta para a reprodução e maturação de uma espécie, sendo estudada em L. vannamei(14), L. setiferus(15), Penaeus monodon(7,16,17), Farfantepenaeus paulensis(18) e P. merguiensis(19). Em camarões de água doce, os estudos foram conduzidos por Samuel et al.(11) com M. malcolmsonii, e Pérez-Rodriguez et al.(20), com M. americanum.
O manejo alimentar para maturação de camarões inclui alimentos frescos, tais como lulas, poliquetas, bivalves, caranguejos e peixes, com ou sem a adição de rações comerciais(6,21). Devido ao risco de transmissão de doenças, tecidos de crustáceos têm sido retirados das dietas de camarões em cativeiro, garantindo desta forma a biossegurança dos reprodutores e não influenciando negativamente no desempenho reprodutivo(16).
De acordo com Harrison(22) e Wouters et al.(23), a importância desses ingredientes frescos dá-se em virtude de seus perfis nutricionais, em particular seus conteúdos e a proporção de certos aminoácidos, frações lipídicas e ácidos graxos essenciais, tais como araquidônico (ARA), eicosapentaenóico (EPA) e docosahexaenóico (DHA), que são de fácil digestão para os crustáceos e reconhecidos por terem significativos papéis metabólicos e fisiológicos na reprodução de peneídeos(24). González-Baró e Pollero(25) relataram a incapacidade dos camarões M. borelli sintetizarem ARA e EPA a partir de outros ácidos graxos, justificando a necessidade de uma suplementação na dieta.
Nesse contexto, o presente estudo teve como objetivo determinar o efeito de três dietas sobre a produção espermática em M. acanthurus.
Material e métodos
Um total de 24 machos adultos de M. acanthurus foram coletados em janeiro de 2014 no rio Sahy (22° 56'S, 44° 01'W) e conduzidos vivos à Estação de Biologia Marinha da Universidade Federal Rural do Rio de Janeiro.
Os camarões foram individualizados em aquários com capacidade de 42 L, dotados de aeração constante, substrato arenoso e abrigos, sendo medidos quanto ao seu comprimento do cefalotórax com auxílio de um paquímetro digital (0,01 mm) e pesados com auxílio de balança analítica com precisão de 0,1 mg, bem como os espermatóforos. A renovação da água ocorreu a cada três dias e parâmetros como temperatura (23,1 ± 1,4°C), pH (7,0 ± 0,5) e oxigênio dissolvido (7,4 ± 0,5 mg/L) foram monitorados diariamente com auxílio de equipamento multiparamétrico (Akso Produtos Eletrônicos, model AK 88); e a amônia (0,9 ± 1,0 ppm) e o nitrito (0,2 ± 0,4 ppm) medidos a cada dois dias (Hanna Instruments Brazil, model HI 83203).
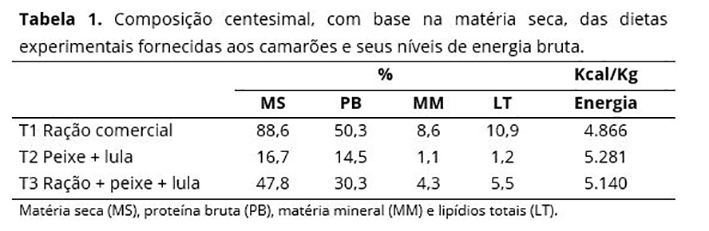
Os animais foram distribuídos em três tratamentos com oito repetições, cada indivíduo constituindo uma unidade experimental, buscando a homogeneidade entre pesos e comprimentos. Cada tratamento foi constituído por uma dieta, sendo compostos por ração comercial (alimento seco) para reprodutores de camarão marinho e dois alimentos frescos, a lula Loligo sp. e o músculo de corvina Micropogonias furnieri. No tratamento T1 foi ofertada apenas ração. Para o tratamento T2, a lula e o músculo do pescado foram adicionados na proporção de 1:1 e processados em moedor de carne até que se obtivesse uma massa homogênea. Na composição da dieta do tratamento T3 foi utilizada uma parte da ração triturada e duas partes da massa obtida com os alimentos frescos.
Após o preparo dos alimentos, uma amostra de 100g de cada dieta foi retirada e enviada ao Laboratório de Bromatologia da UFRRJ para análise de matéria seca (%MS), proteína bruta (%PB), lipídios totais (%LT) e teor de minerais (%MM)(26), cujos resultados estão apresentados na Tabela 1. Para obtenção da energia bruta (Kcal/Kg) foi utilizado 0,5g da amostra previamente processada, analisada em bomba calorimétrica IKA, modelo 2000, isoperibólica, com precisão de 0,001°C(27).
A alimentação foi fornecida diariamente às 10 e 17 h, na base de 10% do peso vivo. A primeira oferta de alimentos só era feita após a extração dos primeiros espermatóforos, via eletroestimulação (6,0 volts), realizada 24 h após o acondicionamento dos animais. Os primeiros materiais seminais compuseram o controle, que continham espermatozoides produzidos no ambiente natural. As demais extrações ocorreram a cada 15 dias, totalizando quatro às realizadas após o início do fornecimento da alimentação em cativeiro, quando os camarões também foram pesados; os mesmos só foram novamente mensurados ao final do período experimental (60 dias). Os espermatóforos foram extraídos de todos os animais, entretanto, apenas os de seis camarões foram utilizados para a contagem da sobrevivência espermática, com o uso de esfregaço de sêmen corado com eosina-nigrosina, cada um constituindo uma unidade experimental.
Antes de cada procedimento de extração, os seis animais que forneciam espermatóforos para a análise da sobrevivência espermática foram sorteados, cabendo aos demais apenas a extração para que todos mantivessem a mesma condição experimental. Somente um espermatóforo por camarão foi aproveitado e quando ocorria a extração de dois, um era escolhido ao acaso. Os outros dois camarões que não compuseram uma rodada de extração, obrigatoriamente cediam material seminal para a extração subsequente, de forma com que todos os animais contribuíssem para a contagem de sobrevivência espermática.
Os dados foram submetidos ao teste Shapiro-wilk para avaliação da normalidade. Para verificar se houve diferença significativa foi aplicada a análise de variância (Anova On-Way) e teste de Tukey, a 5% de significância.
Resultados
Os camarões apresentaram um crescimento de 3,9% em relação ao comprimento do cefalotórax, no tratamento com ração (24,0 ± 4,6 mm inicial; 25,0 ± 3,8 mm final); para os animais do tratamento que consumiram uma mescla de peixe com lula, a taxa de crescimento foi de 0,9% (23,5 ± 2,9 mm inicial; 23,7 ± 2,7 mm final) e para os camarões do tratamento que consumiram uma mescla de alimento fresco com ração foi de 8,5% (22,1 ± 2,2 mm inicial; 24,0 ± 2,2 mm final). Entretanto, a taxa de crescimento não diferiu significativamente entre e dentre os tratamentos.
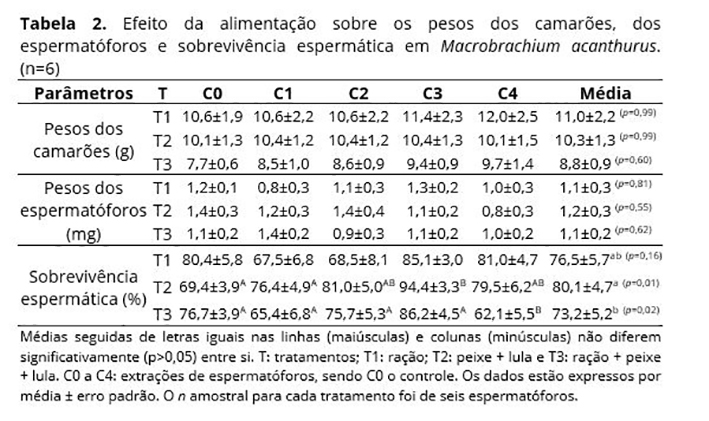
Na Tabela 2 encontra-se o efeito da alimentação sobre os parâmetros biométricos aferidos ao longo do período experimental. Em relação ao peso dos camarões e ao peso dos espermatóforos, não foi verificada diferença significativa entre e dentre os tratamentos durante as diversas coletas realizadas, embora o peso dos camarões alimentados com a mescla de ração com os alimentos frescos tenha se apresentado maior a cada período de extração.
Em relação à sobrevivência espermática, o tratamento T2 foi aquele que apresentou o melhor resultado, obtendo uma média de 80,1%, apesar de não diferir significativamente de T1 (Tabela 2). Entretanto, se considerarmos apenas as quatro coletas em cativeiro, o aumento da sobrevivência foi de 19,4% (T2) em relação ao controle, enquanto os demais tratamentos obtiveram um decréscimo de 6,1% (T1) e 5,7% (T3).
Em relação à produção espermática, 100% dos animais produziram dois espermatóforos durante as coletas. Somente foi observada mortalidade ao final do período experimental, sendo 25% (dois camarões) para cada tratamento.
Discussão
Apesar de M. acanthurus apresentar uma reprodução contínua, de acordo com Tamburus et al.(28), seus picos reprodutivos ocorrem entre dezembro e janeiro na região Sudeste do Brasil. O referido período pode ser considerado ideal para estudos com reprodutores, uma vez que, de acordo com Browdy(6) e Ogle(29), em condições naturais, os fatores ambientais é que determinam a existência de épocas reprodutivas definidas, estimulando ou inibindo o processo reprodutivo de uma espécie em particular, visto que a maturação gonadal está sob controle hormonal, que por sua vez é controlada pelos fatores ambientais.
As temperaturas das águas em ambientes de cultivo são sempre superiores às do ambiente natural e essas maiores temperaturas estariam afetando a sobrevivência espermática, que, independente deste fator, já variam fortemente entre camarões de uma mesma população(30-32).
A média de temperatura encontrada no presente estudo está de acordo com a amplitude térmica determinada por Díaz et al.(33), que mencionaram uma ampla tolerância (15 a 38°C) para M. acanthurus, indicando sua capacidade de se adaptar às diferentes regiões(3), suportando as condições impostas pelo meio sem que comprometessem a produção espermática, uma vez que são nesses intervalos de temperaturas que os animais estão expostos a um mínimo de estresse térmico. Desta forma, as variações térmicas impostas pelo meio aos animais durante o período experimental não comprometeram o processo de produção espermática.
O consumo de oxigênio em M. acanthurus é inversamente proporcional ao peso do animal e maiores animais necessitam de menores concentrações de oxigênio na água(34,35). Segundo Alves et al.(36), o ideal é que a concentração de oxigênio seja superior a 5 mg/L, corroborando os valores reportados no presente estudo, bem como os teores de amônio e nitrito (<1,5 ppm). A baixa qualidade da água, o que não foi constatado no presente estudo, foi reportada como sendo um fator que pode interferir na reprodução(32,37).
Sabe-se que em crustáceos o crescimento é contínuo, mesmo nos animais em período reprodutivo, embora num ritmo mais lento do que nas primeiras fases da vida(38). Entretanto, os ácidos graxos também são responsáveis pelo crescimento em camarões de água doce(39). Papa(40) mencionou em seus estudos com machos de M. amazonicum que dietas contendo maiores quantidades de colesterol inibiram o crescimento corpóreo e testicular do camarão, conforme relatado para outras espécies de decápodos, o que poderia vir a justificar os resultados reportados para o T2, no qual os animais tiveram o menor crescimento (0,9%) em peso/tamanho, apesar da maior sobrevivência espermática (19,4%). Ribeiro et al.(41) mencionaram que os níveis de lipídios podem ser baixos em uma dieta se eles fornecerem uma quantidade suficiente de ácidos graxos, sendo o requerimento destes distinto entre as espécies de crustáceos(25). Deste modo, Leelatanawit et al.(17), quando alimentaram P. monodon com poliquetos, ricos em ácidos graxos (ARA, EPA e DHA), encontraram um aumento de peso e tamanho do camarão a partir da terceira semana, bem como do peso do espermatóforo.
Os lipídios são considerados importantes fontes de energia dos crustáceos. Entretanto, dietas contendo altos níveis desse composto podem não conter quantidades de carboidratos como fonte de glicose, que é ainda mais necessária quando ocorre, por exemplo, síntese de quitina, justificando a presença de uma dieta equilibrada(42). Pérez-Rodríguez et al.(20) relataram que o carboidrato presente na ração seca pode ter sido fundamental para a recuperação da produção espermática em seus animais, que estiveram em cativeiro por um longo período de tempo.
Além de sua importância para a produção de quitina, os carboidratos, bem como as glicoproteínas, estão presentes no fluido seminal, sendo essenciais para o processo de maturação e manutenção da viabilidade espermática(43-46). Jeyalectumie e Subramonian(44) mencionaram a redução dos níveis de quitina em espermatozóides de Scylla serrata preservados a -4°C, indicando seu consumo mesmo a baixas temperaturas. Desta forma, baixas concentrações desse fluido poderiam comprometer a sobrevivência espermática durante os processos de fecundação ou criopreservação, uma vez que não auxiliariam contra as injúrias provocadas pelo meio externo, bem como no fornecimento de energia aos espermatozoides.
Estudos anteriores com camarões marinhos demonstraram que o peso do espermatóforo não depende da quantidade de espermatozoides nele contido(12), mas pode ser associado com a idade do camarão, peso do organismo e dos componentes estruturais(10,20). Assim, o menor peso dos espermatóforos encontrados no T2 poderia estar relacionado a uma carência de carboidratos na dieta fornecida, na base de peixe e lula, embora não tenham sido encontradas diferenças significativas em relação aos demais tratamentos. De acordo com Sacristán et al.(47), os crustáceos decápodes apresentam uma grande diversidade de estratégias de mobilização de suas reservas e ele sugere ainda que as mesmas não estão relacionadas aos tipos de alimento. Pérez-Rodríguez et al.(20) encontraram resultados semelhantes ao do presente estudo, com M. americanum, justificando que os machos necessitam de uma quantidade insignificante de energia para produção de espermatozoides, quando comparado às fêmeas na produção de óvulos.
Shailender et al.(8) e Memon et al.(19) mencionaram que a inclusão de alimentos frescos durante o processo de maturação dos machos é importante para reduzir os efeitos da degeneração natural do espermatóforo, conforme já relatado em outros peneídeos. Além desse fator, segundo Braga et al.(18), F. paulensis alimentados somente com dietas comerciais apresentaram maior ausência de material seminal e hiperglicemia na hemolinfa, não sendo portanto recomendadas, sendo consideradas ideais dietas compostas por um mix entre alimentos fresco e seco.
Em função dos diversos estudos realizados com camarões do gênero Macrobrachium, é sabido que os animais adultos são preferencialmente carnívoros e detritívoros e que aceitam todo tipo de alimento com certo conteúdo de proteína animal, podendo ser alimentados em cativeiro com rações comerciais(48,49). Entretanto, quando apenas estas são ofertadas durante o período de maturação, os resultados da produção, quando comparados aos do alimento fresco, não são satisfatórios, sendo indicado seu uso em um máximo de 50% nas dietas para reprodutores de peneídeos(7,14,50,51). O mesmo foi relatado por Samuel et al.(11) para machos de M. malcolmsonii, que sugeriram o uso de alimento fresco e dietas comerciais.
No presente estudo, a dieta T3 contendo ração comercial e alimento fresco (1:2) não apresentou resultados satisfatórios quanto à sobrevivência espermática, quando comparado ao T2. Entretanto, Perez-Velazquez et al.(51) sugeriram que pelo menos 25% da dieta de maturação deve ser substituída por uma ração, visando conter uma oferta de vitaminas e minerais(9), uma vez que esses nutrientes aparentemente evitam o catabolismo de ARA e EPA, conforme reportado em M. borelli(25).
Quando os animais foram alimentados com 100% de alimento fresco houve perda de peso em L. vannamei e um decréscimo na contagem espermática(14,51), corroborando os resultados do presente estudo. Entretanto, maiores investigações são necessárias para desvendar o papel específico de cada nutriente na reprodução do macho.
Entre os compostos orgânicos alimentares tem sido sugerido que a proteína está mais associada à qualidade de espermatóforos e espermatozoides que os lipídios ou carboidratos, uma vez que a proteína está relacionada com a formação do espinho dos espermatozoides, essencial durante o processo de fertilização dos Dendrobranchiata(15,52,53).
Em seus estudos com machos de L. setiferus, Goimier et al.(15) sugeriram que a proteína dietética em excesso pode desencadear um estresse no animal, provocando a perda da qualidade espermática, assim como o manejo em cativeiro(54). Os maiores teores de proteína aumentam a concentração de hemócitos no camarão, preparando-o para uma defesa imunológica; desta forma, se o animal for estressado por uma quantidade de amônia interna, uma reação imunológica pode ser ativada como resposta ao estresse, afetando várias funções fisiológicas, incluindo a produção e a qualidade dos espermas, seguindo o mesmo mecanismo descrito quando o camarão é submetido a temperaturas extremas(15,37). Entretanto, estes fatores parecem não ter interferido em M. acanthurus, sendo o mesmo reportado por Memon et al.(19), visto que a produção de espermatóforo se manteve ao longo do período experimental, sem apresentar diferença significativa em relação ao controle para a dieta com o maior teor de proteína.
Conclusões
A combinação de uma dieta inerte com uma dieta fresca proporcionou um maior crescimento relativo aos animais e não alterou significativamente o peso dos espermatóforos. Em relação à sobrevivência espermática, a dieta fresca apresentou resultado significativo em relação àquela obtida diretamente do ambiente natural. Entretanto, ressalta-se a necessidade de maiores estudos em relação aos efeitos nutricionais sobre a produção espermática e suas implicações nos processos reprodutivos para camarões de água doce. O presente estudo pode ainda indicar a rusticidade dessa espécie nativa, apresentando boa produção de espermatóforos, mesmo sem uma dieta específica.
Agradecimentos
Agradecemos a todos aqueles que contribuíram para a elaboração deste artigo e à Fundação de Amparo à Pesquisa do Estado do Amazonas – FAPEAM, que financiou este projeto mediante aprovação da bolsa de doutorado (Processo 2801/2011) e a CAPES pelo financiamento da bolsa de doutorado sanduíche no Centro de Investigaciones Biológicas del Noroeste, S.C./B.C.S./México (Processo BEX 4919/14-9).
Referências
1. Camacho ME, Álvarez F, Villalobos JL. Palaemonidae (camarones de río, langostinos). In: Soriano, E.G.; Dirzo, R. e Evogt, R.C. Natural History of the Tuxtlas. Instituto de Biología-UNAM, Instituto de Ecología-UNAM, CONABIO, México; 1997. p. 411-414. English.
2. Coelho PA, Ramos-Porto M. Malacostraca – Eucarida – Caridae (Alpheoidea Excluded). In: Young, PS. Catalogue of Crustacea of Brazil. Museu Nacional, Rio de Janeiro, Brazil. 1998. p. 325-350. Portuguese.
3. Torati L, De Grave S, Page T, Anker A. Atyidae and Palaemonidae (Crustacea: Decapoda: Caridea) of Bocas del Toro, Panama. Check List. 2011;7(6):798-805. English.
4. Valenti WC, Mello JTC, Lobão VL. The fecundity in Macrobrachium acanthurus (Wiegmann, 1836) of the Ribeira de Iguape river (Crustacea, Decapoda, Palaemonidae). Revista Brasileira de Zoologia. 1989;6(1):9-15. Portuguese.
5. Arcos-Ortega GF. Physiological and genetic analysis of reproductive performance of white shrimp Litopenaeus vannamei. [Tese]. La Paz, Baja California Sur, México: Centro de Investigaciones Biológicas del Noroeste; 2004. 244 p. [Acesso 2013 Out. 22] Disponível em: http://www. oceandocs.org/bitstream/handle/1834/3524/arcos_g.pdf?sequence=1. Espanish.
6. Browdy CL. A review of reproductive biology of Penaeus species: Perspectives on controlled shrimp maturation systems for high quality nauplii production. In: Wyban J. Proceedings of the special session on shrimp farming. World Aquaculture Society, Baton Rouge, Los Angeles, USA. 1992. p. 22-51.
7. Meunpol O, Meejing P, Piyatiratitivorakul S. Maturation diet based on fatty acid content for male Penaeus monodon (Fabricius) broodstock. Aquaculture Research. 2005;36(1):1216-1225. English.
8. Shailender M, Suresh Babu CH, Krishna PV. Determine the competence of different fresh diets to improve the spermatophore superiority of giant black tiger shrimp, Penaeus monodon (Fabricius, 1798). International Journal of Bioassays. 2012;1(12):170-176. English.
9. Bray WA, Lawrence AL. Reproduction of Penaeus species in captivity. In: Fast AW, Lester LJ. Marine Shrimp Culture: principles and practices. Elsevier, New York, USA. 1992. p. 93-171. English.
10. Alfaro-Montoya J, Lozano X. Development and deterioration of spermatophores in pond-reared Penaeus vannamei. Journal of the World Aquaculture Society. 1993;24(4):522-529. English.
11. Samuel MJ, Kannupandi TY, Soundarapandian P. Nutritional effects on male reproductive performance in the freshwater prawn Macrobrachium malcolmsonii (H. Milne Edwards). Aquaculture. 1999;172(1):327-333. English.
12. Díaz AC, Fernandez-Gimenez AV, Harán NS, Fenucci JP. Reproductive performance of male Argentine red shrimp Pleoticus muelleri Bate (Decapoda, Penaeoidea) in culture conditions. Journal of the World Aquaculture Society. 2001;32(2):236-242. English.
13. Harlioğlu MM, Farhadi A, Gür, S. Determination of sperm quality in decapod crustaceans. Aquaculture. 2018;490(1):185-193. English.
14. Perez-Velazquez M, Lawrence AL, Gatlin DM, González-Félix ML, Bray WA. Replacement of fresh dietary components by a dry feed for successful maturation of male Litopenaeus vannamei (Boone) broodstock. Aquaculture Research. 2002;33(1):1901-1905. English.
15. Goimier Y, Pascual C, Sánchez A, Gaxiola G, Sánchez A, Rosas C. Relation between reproductive, physiological, and immunological condition of Litopenaeus setiferus pre-adult males fed different dietary protein levels (Crustacea, Penaeidae). Animal Reproduction Science. 2006;92(1): 193-208. English.
16. Coman GJ, Arnold SJ, Callaghan TR, Preston NP. Effect of two maturation diet combinations on reproductive performance of domesticated Penaeus monodon. Aquaculture. 2007;263(1):75-83. English.
17. Leelatanawit R, Uawisetwathana U, Khudet J, Klanchui A, Phomklad S, Wongtripop S, Angthoung P, Jiravanichpaisal P, Karoonuthaisiri N. Effects of polychaetes (Perinereis nuntia) on sperm performance of the domesticated black tiger shrimp (Penaeus monodon). Aquaculture. 2014;433(1):266-275. English.
18. Braga AL, Nakayama CL, Martins JG, Colares EP, Wasielesky Jr. W. Spermatophore quality of the pink shrimp Farfantepenaeus paulensis (Decapoda, Dendrobranchiata) broodstock fed with different maturation diets. Aquaculture. 2010;307(1):44-48. English.
19. Memon AJ, Ikhwanuddin M, Talpur AD, Khan MI, Fariddudin MO, Safiah J, Abol-Munafi AB. To determine the efficiency of different fresh diets in improving the spermatophore quality of banana shrimp Penaeus merguiensis (De Man, 1888). Journal of Animal and Veterinary Advances. 2012;11(9):1478-1485. English.
20. Pérez-Rodríguez JC, Gómez-Gutiérrez J, López-Greco LS, Cortés-Jacinto E. Spermatophore production and sperm quality of the river prawn Macrobrachium americanum Spence Bate, 1868 fed with different diets. Aquaculture Research. 2019;50(1):3117-3129. English.
21. Harrison K. Broodstock nutrition and maturation diets. In: D'Abramo LR, Conklin DE, Akiyama DM. Crustacean nutrition. Advances in World Aquaculture. The World Aquaculture Society, Baton Rouge, USA. 1997. p. 390-408. English
22. Harrison, K. The role of nutrition in maturation, reproduction and embryonic development of decapod crustaceans. A review. Journal of Shellfish Research. 1990;9(1):1-28. English.
23. Wouters R, Lavens P, Nieto J, Sorgeloos P. Penaeid shrimp broodstock nutrition: an updated review on research and development. Aquaculture. 2001;202(1):1-21. English.
24. Glencross BD. Exploring the nutritional demand for essential fatty acids by aquaculture species. Reviews in Aquaculture. 2009;1(1):71-124. English.
25. González-Baró MR, Pollero RJ. Fatty acid metabolism of Macrobrachium borelli: dietary origin of arachidonic acid and eicosapentaenoic acid. Comparative Biochemistry and Physiology - Part A. 1998;119(3):747-752. English.
26. A.O.A.C. Official methods of analysis. 17ª ed. Association of Official Analytical Chemists, Gaithersburg, Maryland, USA. 2000. 2200p. English.
27. Ika Werke. Ika calorimeter system C 2000 operating instructions. USA. 2007. Portuguese.
28. Tamburus AF, Mossolin EC, Mantelatto FL. Populational and reproductive aspects of Macrobrachium acanthurus (Wiegmann, 1836) (crustacea: palaemonidae) from north coast of São Paulo state, Brazil. Brazilian Journal of Aquatic Science and Technology. 2012;16(1):9-18. English.
29. Ogle JT. A review of the current (1992) state of our knowledge concerning reproduction in open thelycum Penaeid shrimp with emphasis on Penaeus vannamei. Invertebrate Reproduction and Development. 1992;22(1-3):267-274. English.
30. Bray WA, Leung-Trujlllo JR, Lawrence AL, Robertson SM. Preliminary investigation of the effects of the temperature, bacterial inoculation, and EDTA on sperm quality in captive Penaeus setiferus. Journal of the World Mariculture Society. 1985;16(1-4):250-257. English.
31. Pascual C, Valera E, Re-Regis C, Gaxiola G, Sanchez A, Ramos L, Soto LA, Rosas C. Effect of water temperature on reproductive tract condition of Penaeus setiferus adult males. Journal of the World Aquaculture Society. 1998;29(4):477-484. English.
32. Perez-Velazquez M, Bray WA, Lawrence AL, Gatlin DM, González-Félix ML. Effect of temperature on sperm quality of captive Litopenaeus vannamei broodstock. Aquaculture. 2001;198(1):209-218. English.
33. Díaz F, Sierra E, Re AD, Rodríguez L. Behavioral thermoregulation and critical thermal limits of Macrobrachium acanthurus (Wiegmann). Journal of Thermal Biology. 2002;27(1):423-428. English.
34. Elmor MRD, Lobão VL, Valenti WC. Oxygen-consumption by the shrimp Macrobrachium acanthurus (Wiegmann, 1836) as subsidy to its transport and cultivation. Boletim do Instituto de Pesca. 1981;8(único):65-78. English.
35. Gasca-Leyva JFE, Martinez-Palacios CA, Ross LG. The respiratory requeriments of Macrobrachium acanthurus (Wiegman) at different temperatures and salinities. Aquaculture. 1991;93(1):191-197. English.
36. Alves PC, Ramos-Porto M, Soares CMA. Cultivation of Macrobrachium bate genus shrimp (Decapoda, Palaemonidae) from Brazil. Boletim Técnico 6, EMPARN, Natal, Rio Grande do Norte, Brazil. 1981. 66p. Portuguese.
37. Pascual C, Sánchez A, Sánchez A, Vargas-Albores F, Lemoullac G, Rosas C. Haemolymph metabolic variables and immune response in Litopenaeus setiferus adult males: the effect of an extreme temperature. Aquaculture. 2003;218(1):637-650. English.
38. Albertoni EF, Palma-Silva C, Esteves FA. Distribution and growth in adults of Macrobrachium acanthurus Wiegmann (Decapoda, Palaemonidae) in a tropical coastal lagoon, Brazil. Revista Brasileira de Zoologia. 2002;19(Supl.2):61-70. English.
39. D'Abramo LR, Sheen SS. Polyunsaturated fatty acid nutrition in juvenile freshwater prawn Macrobrachium rosenbergii. Aquaculture. 1993;114(1):63-86. English.
40. Papa LP. Structural aspects of the testes of different morphotypes of Macrobrachium amazonicum hand fed diets with different levels of cholesterol. [Dissertação]. Jaboticabal, São Paulo, Brazil: Universidade Estadual Paulista, Caunesp. 2003. 81f. [Acesso 2014 Nov. 13]. Disponível em: http://repositorio.unesp.br/bitstream/handle/11449/144106/000215426.pdf?sequence=1&isAllowed=y. Portuguese.
41. Ribeiro K, Franceschini-Vicentini IB, Papa LP, New MB, Valenti WC. Effect of polyunsaturated fatty acids on the fecundity of the Amazon river praw Macrobrachium amazonicum (Heller, 1862). Aquaculture Research. 2012;43(1):1756-1763. English.
42. Gastelú JC, Oliveira J, Brito LO, Galvez AO, Moreira MG. Effect of temperature and food protein and lipid in stages of ovarian maturation and stages of molting Macrobrachium acanthurus (Wiegmann, 1836). Ciência Animal Brasileira. 2011;12(3):443-455. Portuguese.
43. Dougherty WJ, Dougherty MM, Harris SG. Ultrastructural and histochemically observations on eletroejaculated spermatophores of the palaemonid shrimp Macrobrachium rosenbergii. Tissue & Cell. 1986;18(5):709-724. English.
44. Jeyalactumie C, Subramoniam T. Cryopreservation of spermatophores and seminal plasma of the edible crab Scylla serrata. Biological Bulletin. 1989;177(1):247-253. English.
45. Poljaroen J, Vanichviriyakit R, Tinikul Y, Phoungpetchara I, Linthong V, Weerachatyanukul W, Sobhon P. Spermatogenesis and distinctive mature sperm in the giant freshwater prawn, Macrobrachium rosenbergii (de Man, 1879). Zoologischer Anzeiger. 2010;249(1):81-94. English.
46. Zara FJ, Toyama MH, Caetano FH, López-Greco LS. Spermatogenesis, spermatophore, and seminal fluid production in the adult blue crab Callinectes danae (Portunidae). Journal of Crustacean Biology. 2012;32(2):249-262. English.
47. Sacristán HJ, Rodríguez YE, Pereira NA, López-Greco LS, Lovrich GA, Gimenez AVF. Energy reserves mobilization: Strategies of three decapod species. PLoS ONE. 2017;12(9):1-19. English.
48. Albertoni EF, Palma-Silva C, Esteves FA. Natural diet of three species of shrimp in a Tropical Coastal Lagoon. Brazilian Archives of Biology and Technology. 2003;46(3):395-403. English.
49. García-Guerrero MU, Becerril-Morales F, Vega-Villasante F, Espinosa-Chaurand LD. The Macrobrachium prawns with economic and fisheries importance in Latin America: present knowledge, ecological role, and conservation. Latin American Journal of Aquaculture Research. 2013;41(4):651-675. English.
50. Wouters R, Zambrano B, Espin M, Calderon J, Lavens P, Sorgeloos P. Experimental broodstock diets as partial fresh food substitutes in white shrimp Litopenaeus vannamei B. Aquaculture Nutrition. 2002;8(1):249-256. English.
51. Perez-Velazquez M, González-Félix ML, Lawrence AL, Bray WA, Gatlin DM. Dietary effects on sperm quality of Litopenaeus vannamei broodstock. Journal of the World Aquaculture Society. 2003;34(1):92-98. English.
52. Sánchez A, Pascual C, Sanchez A, Vargas-Albores F, Moullac GL, Rosas C. Hemolymph metabolic variables and immune response in Litopenaeus setiferus adult males: the effect of acclimation. Aquaculture. 2001;198(1):13-28. English.
53. Braga AL, Nakayama CL, Poersch LH, Wasielesky Jr. W. Unistellate spermatozoa of decapods: comparative evaluation and evolution of the morphology. Zoomorphology. 2013;132(1):261-284. English.
54. Leung-Trujillo JR, Lawrence AL. Observations on the decline in sperm quality of Penaeus setiferus under laboratory conditions. Aquaculture. 1987;3-4(1):363-370. English.