
Abstract
Native bees, besides excellent producers of honey, lend a great legacy to humanity through their ability to pollinate plants and the consequent preservation of ecosystems. In this interspecific relation, bees are benefited by plant species by the provision of food (nectar and pollen), besides using their branches and trunks for lodging. However, the anthropogenic action has jeopardized the survival of stingless bees, causing irreversible environmental damage in the preservation of natural resources. This study aimed to identify the nesting habits of native bees under natural conditions. Random trails in the Curimataú micro-region of Paraíba allowed identifying nesting sites of indigenous species in their natural habitat. Sixty honeycomb nests were identified in 12 different plant species, being Commiphora leptophloeos the most preferred host (46.66%), with 55% of the entrance holes pointed to magnetic orientations between the Northeast and Northwest; a greater choice by host trees (85%); preference for nesting in host plants with a circumference of 0.98 m in diameter, entrance hole of nests of 7.77 mm in diameter, and height from the ground of 1.52 m. The obtained data showed little variety of stingless bee species in the Curimataú region of Paraíba, especially Melipona subnitida. The species Melipona asilvae, Melipona scutellaris, Partamona seridoenses, and Plebeia sp. should receive special attention not to become extinct in this region.
Keywords: Curimataú. Natural habitat. Nesting. Preference. Stingless bees.
Resumo
As melíponas, além de excelentes produtoras de mel, prestam um grande serviço à humanidade através da sua capacidade de polinizar plantas e sua consequente preservação de ecossistemas. Nessa relação interespecífica harmônica, as abelhas são beneficiadas pelas espécies vegetais pelo provimento do alimento (néctar e pólen), além de utilizarem seus galhos e troncos para se alojarem. Porém, a ação antrópica predatória tem colocado em risco a sobrevivência das melíponas, causando danos ambientais irreversíveis e comprometendo a preservação dos recursos naturais. Este trabalho objetivou identificar os hábitos de nidificação das abelhas nativas em condições naturais. Por meio de trilhas aleatórias na microrregião do Curimataú paraibano foi possível identificar os pontos de nidificação das espécies de meliponíneos em seu habitat natural. Foram identificados 60 ninhos de melíponas nidificando em 12 espécies vegetais diferentes, sendo a Commiphora leptophloeos a hospedeira de maior preferência (46,66%), com 55% dos orifícios de entrada apontados para orientações magnéticas entre o Nordeste e Noroeste; com maior escolha por hospedeiras (árvores) [85%]; apresentando preferência por nidificação em vegetais hospedeiros com circunferência de 0,98 m de diâmetro, orifício de entrada dos ninhos com 7,77 mm de diâmetro e uma altura do solo de 1,52 m; Os dados obtidos revelam pouca variedade de espécies melíponas na região Curimataú paraibano, com destaque para Melipona subnitida. As espécies Melipona asilvae, Melipona scutellaris, Partomona seridoenses e Plebeia sp. devem ter uma atenção especial para não entrarem em extinção nessa região.
Palavras-chave: Curimataú. Habitat natural. Melíponas. Nidificação. Preferência.
Section: Zootecnia
Received
May 25, 2019.
Accepted
June 18, 2019.
Published
August 6, 2020.
www.revistas.ufg.br/vet
visit the website to get the how to cite in the article page.
Introduction
Meliponines, also known as stingless or indigenous bees, inhabit the Earth about 125 million years.(1) Meliponiculture is an ancient activity that has been practiced since the dawn of civilizations in ancient Egypt.(2) In Brazil, the rearing of honey bees was initiated by indigenous(3) even before the discovery and introduction of Apis mellifera L. (Hymenoptera: Apidae).
Bees provide a great service to humanity through their ability to pollinate, as they are insects specialized in plant pollination,(4) contributing to the preservation of ecosystems and generating an increase in the production of commercial plant species.(5) In addition, they are the main pollinating agents on the planet. Approximately 80% of the world's cultivated plant species are estimated to be pollinated by some bee species, 19% by flies, 6.5% by bats, 5% by wasps, 5% by beetles, 4% by birds, and 4% by butterflies and moths.(6)
The indiscriminate use of pesticides has endangered fauna and flora due to their harmful effects on pollinating insects of agroecosystems.(7) One of the major consequences of this practice is the direct reduction of the bee population, as well as indirect effects of economic losses resulting from the decline of these pollinators.(8)
Anthropogenic action, i.e., the predatory action of humans over decades has caused irreversible environmental damage in the preservation of natural resources. Advances in agricultural frontiers have increased the rates of deforestation and indiscriminate use of pesticides, making it difficult to fix native bees in their habitats. As a result, stingless bees are in an accelerated process of disappearance, mainly caused by the loss of environmental quality related to the clearing of native forests.(9) Therefore, the degradation and destruction of the environment seem to be the main factor of biodiversity decline.(10) Other factors, however, have been causing concern in the preservation of these species, such as global warming.(11)
An imminent and compromising threat to the future of meliponiculture is attributed to "honey hunters," people who irregularly exploit the trade of indigenous bee honey. These people cut down the trees in search of native bees, rudely remove the honey, destroy the honeycomb frames of beehives, and throw the remaining ones on the ground, which are eaten by ants. It rapidly leads to the decimation of stingless bees(12, 13) by the destruction of their nests through this indiscriminate and predatory action practiced by honey hunters.(14)
The rearing of indigenous bees, often performed empirically by producers, generates a demand for research that contributes to production with appropriate management techniques.(15) This study aimed to identify the nesting habits of stingless bees under natural field conditions regarding their occurrence, location, plant substrates, substrate characteristics, architecture, and nesting preferences, also considering the magnetic orientation in the Curimataú micro-region of Paraíba, besides proposing technical specifications that may be used in the manufacture of standard boxes, orientation of their positioning in relation to the cardinal points, and rational and humanized management of stingless bees in a sustainable way.
Stingless bees are docile bees that live in colonies and are easy to manage.(17) Understanding the nesting process of stingless bees is of fundamental importance for species preservation and careful planning of a sustainable production system, taking into consideration the region of their occurrence and characteristics of the habitat in which they are inserted. Based on this understanding, several authors(18–22) described the preference of different nesting habits for stingless bee species.
The diameter of the tree trunk that house the stingless bee nests gives an idea of the diameter of the pre-existing cavity occupied by the nest and may vary from species to species. Most species depend on these pre-existing cavities, usually in cavities of living or dead trees, to build their nests,(23) and their internal diameter varies with nested species.(24)
The orientation ability of living beings is a determining factor in species survival, whether in displacement, food collection, or reproduction. Thus, the most varied species develop mechanisms of adaptation to the environment. Among these abilities, the theory of the ability to detect the Earth's magnetic field for orientation and navigation, commonly accepted by several researchers,(25–27) is noteworthy, although it is little known in animals.(28)
The substrate is another parameter that undergoes variations regarding nesting habits of stingless bees. Sometimes, several nesting habits are found within the same genus of stingless bees.(22) The factors that drive the preference of bees for a particular plant source are not known,(29) but reports show they are selective in this choice. In general, stingless bees depend on trees for housing and food,(30, 24) building their nests in pre-existing cavities. The region of occurrence of all species of stingless bees must be respected in a rational exploration, avoiding the degradation of that ecosystem. Understanding the pattern of their distribution and abundance is considered important(31) for developing conservation strategies in order to reduce the effects of habitat fragmentation on the genetic and ecological structure of stingless bee populations.
The bees are part of the order Hymenoptera and meliponines are in the superfamily Apoidea, which is subdivided into the families Andrenidae, Anthophoridae, Colletidae, Halictidae, Melittidae, Megachilidae, Oxaeidae, and Apidae. The family Apidae, in its turn, is divided into four subfamilies: Bombinae, Euglossinae, Apinae, and Meliponinae. The subfamily Meliponinae is divided into two tribes: Meliponini and Trigonini. Bees of both tribes can be called meliponines.(32) Fifty-two genera, with a total of 400 species,(33) can be found within the subfamily Meliponinae spread throughout the world, from the state of Rio Grande do Sul in Brazil to central Mexico, Africa, India, Malaysia, Indonesia, and Australia.(32)
Brazil is one of the main places of stingless bee occurrence, where they have a great influence on the maintenance of ecosystems. To date, 33 genera of Neotropical meliponines have been described in Brazil, comprising 300 species,(34) which may vary between 350 and 400 species.(35) However, it is believed that the number of species is much higher, with a high diversity of shapes, size, and nesting habits.(36) At least 100 stingless bee species are in danger of extinction due to the destruction of their habitats by humans.(32)
The tribe Meliponini is unique of the Americas, in the Neotropical region (from southern South America to the mountains of northern Mexico in Central America), with approximately 26 species and numerous subspecies, and is represented by one genus: Melipona. In Brazil, there are about 20 species: Melipona scutellaris, M. mandacaia, M. asilvae, M. quinquefasciata, M. subnitida, among others.(37) In Paraíba, the tribe Meliponini is distributed in the coastal micro-regions, Curimataú, Borborema, and Sertão, with five species: M. subnitida, M. quadrifasciata, M. asilvae, M. compressipes fasciculata, and M. scutellaris.(38)
The tribe Trigonini is a very diverse group of stingless bees, encompassing the largest number of genera in the Neotropical region, being possible to find several genera: Lestrimelitta, Hypotrigona, Cephalotrigona, Oxytrigona, Schwarziana, Scaura, Plebeia, Partamona, Paratrigona, Scaptotrigona. Nannotrigona, Tetragonisca, Trigona, Geotrigona, and Frieseomelitta, and numerous species (around 120).(37) In Paraíba, thirteen Trigonini species were found into eight different genera: Frieseomelitta, Trigona, Scaptotrigona, Partamona, Tetragonisca, Leurotrigona, Plebeia, and Nannotrigona.(38)
Caatinga is a unique biome, located in the semi-arid climate, and is the main existing ecosystem in the Northeast region of Brazil, the natural cradle of stingless bees. This region has a great variety of landscapes with relative biological and endemic richness, governed by seasonal and periodic droughts that are renewed with each rainfall cycle. This biome has been undergoing extensive environmental devastation caused by an anthropogenic process of indiscriminate use of its natural resources.(16)
In Paraiba, Curimataú is a micro-region within the Caatinga biome known for the presence of indigenous bees. Municipalities of this locality, such as Araruna, Cacimba de Dentro, Damião, and Cuité, have a vast predominantly Caatinga area and semi-arid climate, marked by low precipitation, short, poorly distributed rainfall, and with fauna and flora threatened by anthropogenic action. Thus, knowing the nesting habits of stingless bees in this region is essential for meliponiculturists to improve their management techniques. This study aimed to identify the nesting habits of native bees under natural conditions in the Curimataú micro-region of Paraíba.
Material and methods
This study was carried out in the Curimataú region of Paraíba. For sampling purposes, the data were collected from September to November 2016 from four municipalities: Araruna (06°33′30″ S and 35°44′30" W), Damião (06°37′54″ S and 35°54′18″ W), Cuité (06°29′01″ S and 36°09′13″ W), and Cacimba de Dentro (06°38′21″ S and 35°46′42″ W), totaling an area of 1,336.9 km². Nesting sites were previously identified through interviews with family farmers living in the collection areas.
A survey of possible nesting sites of native bees in the Curimataú region of Paraíba started via random trails with the assistance of native informers from the region. The botanical identification of host trees was performed by collaborators using the common name of the species and confirmed by a search in specialized websites (Tópicos and Flora do Brasil).
Nesting sites were cataloged by the Global Navigation Satellite System (GPS) using a Garmin® etrex 30 device, allowing identifying their geographical coordinates and magnetic orientation (cardinal and collateral points) of the entrance hole of nests preferred by native bees. Other parameters were evaluated for better characterization and identification of native bee preference when choosing nesting sites. The methodology described by Serra(31) was used by measuring the diameter of the entrance hole of nests with a Lotus® caliper, tree diameter at the entrance hole location with a Stanley® measuring tape, and the height of the entrance hole of nests in relation to the ground height with a 5-meter Irwin® measuring tape. An entomological net was used for the capture of samples with a hole height higher than 2 meters.
A sample of five native bee specimens was collected at each nesting site by placing them in an acrylic container (50 ml) with a lid containing 30 ml alcohol (70%). The container was duly identified and sent to the Laboratory of Entomology (LE) of the Department of Systematics and Ecology (DES) of the Federal University of Paraíba (UFPB) for taxonomic analysis.
The chi-square test was used for occurrence frequencies of species, while Duncan's test was used after ANOVA to compare means of the diameter of the entrance hole, trunk diameter, and nest height as a function of species and orientation.
Results and discussion
Sixty nests of stingless bees were found in four different municipalities of the Curimataú region of Paraíba (Araruna, Cacimba de Dentro, Damião, and Cuité). Bees (tribes Meliponini and Trigonini) corresponded to eight different species: Melipona subnitida Ducke, 1910, Melipona asilvae Moure, 1971 (Hymenoptera: Apidae), Melipona scutellaris Latreille, 1811 (Hymenoptera: Apidae), Frieseomelitta doederleini Friese, 1900 (Hymenoptera: Apidae), Frieseomelitta francoi Moure, 1946 (Hymenoptera: Apidae), Partamona seridoenses Pedro & Camargo, 2003 (Hymenoptera: Apidae), Plebeia sp., and Scaptotrigona aff. depilis Moure, 1942 (Hymenoptera: Apidae). The most frequent species was M. subnitida (36.67%), followed by F. doederleini (25%), and Frieseomelitta francoi Moure, 1946 (Hymenoptera, Apidae) and S. aff. depilis, both with 11.67% (Table 1). A study carried out in Serra da Capivara, PI,(39) also observed the occurrence of these species.
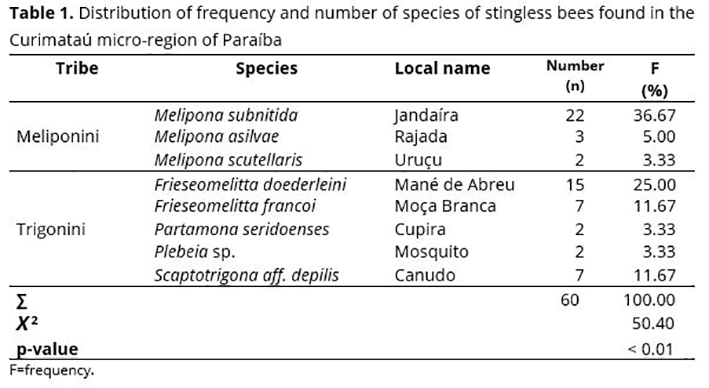
Among the 18 native species of bees that inhabit Paraiba,(38) eight were found distributed in the collection area (Curimataú micro-region of Paraíba), in which the species at higher occurrence in the study area was Melipona subnitida, with 36.67% of the nests. It confirms the predominance of this species in the Curimataú micro-region,(38) followed by Frieseomelitta doederleini (25%) and Scaptotrigona aff. postica Latreille, 1807 (Hymenoptera: Apidae) (11.67%). The bee species Melipona scutellaris, Partamona seridoenses, and P. sp. were the least frequent (all of them with 3.33%) (Table 1). Considering that stingless bee species are under an accelerated process of disappearance(9) caused especially by the loss of habitat due to deforestation of native forests (their natural environment), their low incidence suggests that an environmental education program be carried out in local communities, particularly with meliponiculturists. In a field research carried out in João Câmara, Rio Grande do Norte, Martins et al.(40) found seven species of stingless bees, in which M. subnitida was more frequent (57.3%), followed by the species Melipona asilvai Moure, 1971 (Hymenoptera: Apidae) (17.2%) and Frieseomelitta dispar Moure, 1950 (Hymenoptera: Apidae) (13.7%).
Although the hundreds of tree species in the Curimataú region, stingless bees have used a small number of them for nesting, i.e., only 12 species (Table 2). Stingless bee species are selective and opt for specific trees.(41)
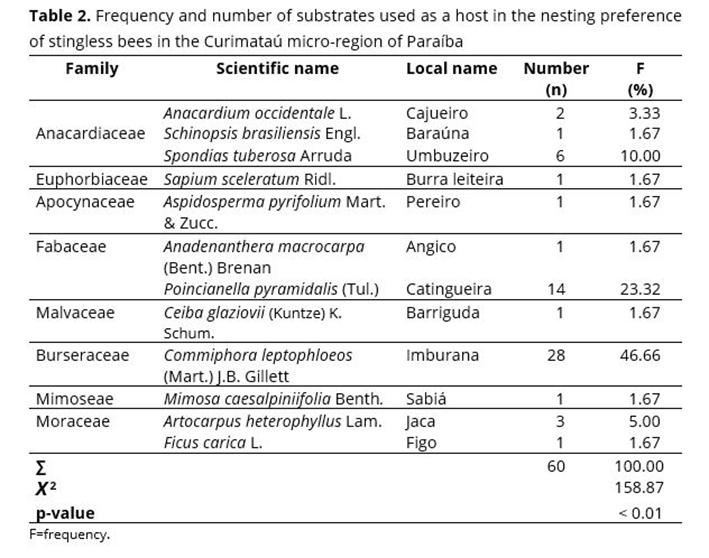
This study showed that stingless bees nest in different tree species. However, 69.98% of the nests were observed in only two tree species: Commiphora leptophloeos (Mart.) J.B. Gillett, with 46.66%, followed by Poincianella pyramidalis (Tul.), with 23.32% (Table 2). Similar data were found in phytosociological studies carried out in a secondary growth Caatinga area in the Seridó region,(42) where Poincianella pyramidalis had the highest densities and frequencies. The species C. leptophloeos has a hollow trunk, an essential condition for nest installation. Similar results in the preference of host plant species with stingless bees in the Caatinga (Seridó, PB, and João Câmara, RN) were observed(40) for C. leptophloeos and Poincianella pyramidalis, with a frequency of 75.0%, which is similar to that found in this study (69.98%).
Knowledge on the preference of stingless bees for nesting choices goes beyond a cavity and good flowering. Magnetic orientation, i.e., the direction the entrance hole of nests is facing, should also be taken into consideration.
Stingless bees found in the Curimataú micro-region of Paraíba have a specific preference in the magnetic orientation. Cardinal and collateral points with variable orientation between Northeast and Northwest (Table 3) had a higher choice, with a frequency of 55%.

The lowest values for magnetic orientation were found for Southeast (3.33%), South (6.67%), and Southwest (6.67%) positions, totaling 16.67% .
In their natural habitat, stingless bees seek to install their nests in camouflaged locations. Most of these bees use cavities in living trees, dead trees, dry trunk, stones, termite mounds, anthills, wall tiles, among others. However, nesting in live tree trunks is predominant.(43)
The results showed that 85% of stingless bees nest in cavities of living trees, followed by dead trees (6.67%), dry trunks (5.0%), and termite mounds (3.33%) (Table 4). A study carried out in Cataguases, MG, showed that 20 of 21 nests were found in a tree substrate, and only one nest of T. angustula was built in a ravine.(44) It is evident the preferences of stingless bees for live trees to nest.
Only 3.33% of the species nested in termite mounds (Table 4). These termite mounds, in turn, were occupied by P. seridoenses, a bee that has a specific nesting habit in termite mounds.(45)
Stingless bees have characteristics that can be influenced at nesting time in relation to the diameter of the entrance hole, trunk diameter, and nest height. Nest hole sizes of the studied stingless bees differed between species (Table 5). However, nest height showed no difference between species.
The diameter of the entrance hole of stingless bee nests may vary when considering the size of each species, i.e., the larger their size is, the larger the entrance hole. Significant differences in the diameter of the entrance hole were observed when comparing the studied species (Table 5). The largest diameter was observed for Scaptotrigona (10.43 mm), with no statistical difference from M. scutellaris (9.0 mm), M. subnitida (8.45 mm), and P. seridoensis (7.50 mm).

Intermediate results for the diameter of the entrance hole were recorded for P. seridoensis (7.5 mm), F. doederleini (7 mm), and F. francoi (6 mm), while the lowest value was found for M. asilvae (4.67 mm) (Table 5). Stingless bees typically build their nests in pre-existing native tree cavities, with an internal diameter varying according to the nested species.(24)
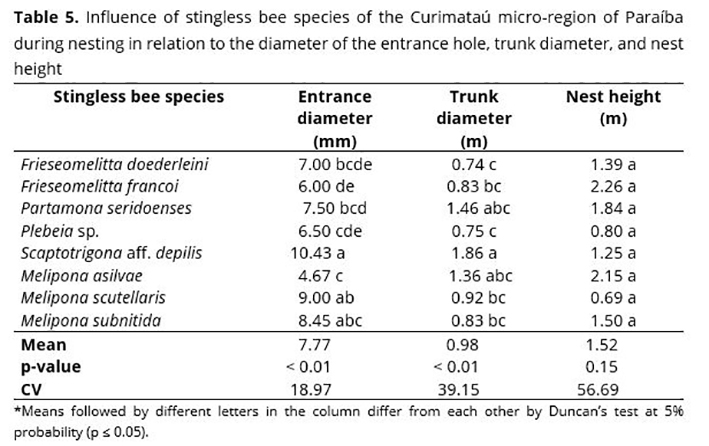
Trunk diameter was influenced by the studied stingless bee species, in which the highest values were found for S. aff. depilis (1.86 m), P. seridoenses (1.46 m), M. asilvae (1.36 m), and M. scutellaris (0.92 m), differing from M. subnitida, which ranged from 0.74 to 0.83 m (Table 5). Other studies have found different results, with diameter values ranging from 0.39 and 0.50 m.(40)
Trunk diameter may be directly related to the occupation volume used by stingless bees, which may vary regarding the species in relation to their requirements to build their nests. The capacity of the pre-existing cavity may be decisive in choosing the nesting site.(46) Beehive volume is a determining factor and may vary according to the region.(32) Usually, an M. subnitida beehive requires a volume of 15 liters.
Nest height did not change significantly when comparing stingless bee species, although they presented a variation of 0.80 m for Plebeia sp. and 2.6 m for F. francoi (Table 5), which led to a high coefficient of variation. A significant influence of the host plant species of the Curimataú micro-region of Paraíba was observed in relation to the diameter of the entrance hole and trunk diameter of nests found in their natural habitat (Table 6). Also, no significant difference was observed for nest height.
The largest entrance holes were found in the species A. heterophyllus and F. carica (11 mm), A. occidentale (10 mm), and C. glaziovii and S. sceleratum (9 mm). The lowest values were observed for A. macrocarpa, A. pyrifolium, and S. brasiliensis (4, 5, and 4 mm, respectively) (Table 6).
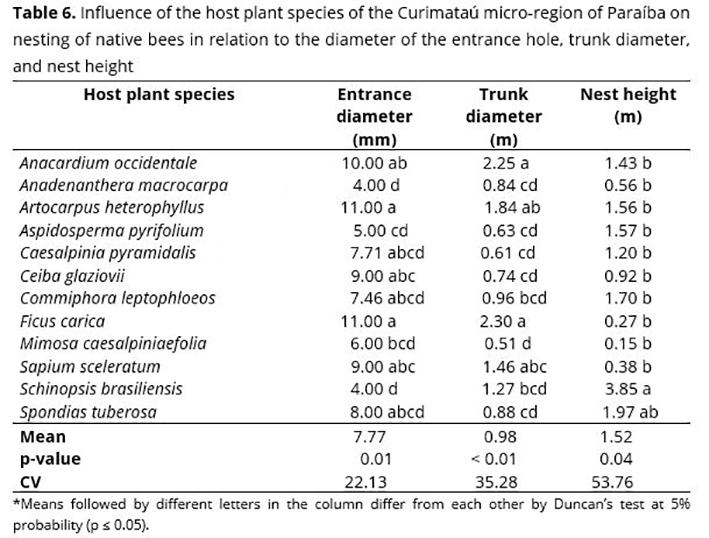
When comparing tree trunk diameter in relation to the influence of host plants (Table 6), it is observed that the largest diameters were presented by A. occidentale and F. carica, with 2.25 and 2.30 m, respectively, differing statistically from M. caesalpiniaefolia, which had the lowest value (0.51 m). A study conducted in the Forest Zone of Pernambuco showed that 50% of stingless bees nest in trees with a diameter lower than 50%, 30% with a diameter between 1 and 1.5 m, and 20% between 2 and 2.5 m.(47) Diameter variation of trees chosen for nesting of stingless bee species found in the Curimataú region of Paraíba may be related to the size of trees in the study area, with varying characteristics regarding size and architecture.
Nest height showed no significant difference regarding host tree species, even though it showed a maximum amplitude of 3.85 m for S. brasiliensis and minimum for M. caesalpiniaefolia, with 0.15 m of nest height in relation to the ground. Similar results were found in a survey conducted in Bahia with stingless bees,(48) in which nest height ranged from 0.5 to 3.8 m. Probably, nest height of stingless bees is not related to the size or type of trees, but rather to nesting characteristics of each bee species. The height of nest entrance hole in host plant species at the highest or lowest positions is due to chance.(31) Cardinal and collateral points showed no influence on the diameter of the entrance hole, trunk diameter, and nest height (Table 7).
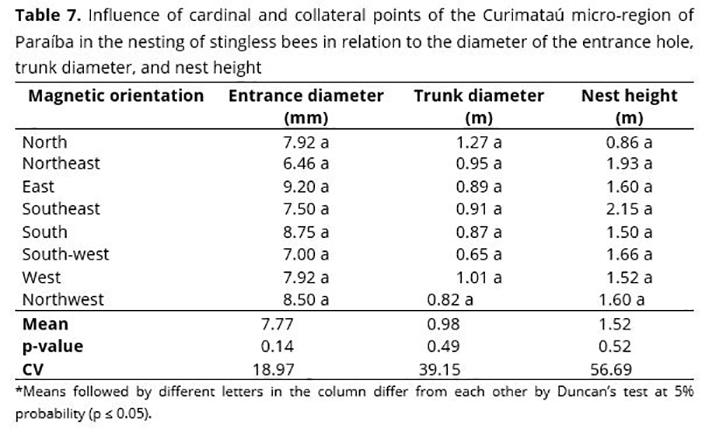
Although not statistically different, the diameter of the entrance hole showed the highest values for points facing East, South, and Northwest, with 9.2, 8.75, and 8.5 mm, respectively. The lowest values were observed for Northeast, Southeast, and Southwest (6.46, 7.5, and 7.0 mm, respectively (Table 7).
Trunk diameter of host trees was not significantly influenced when comparing cardinal and collateral points (Table 7), although the largest diameter was 1.27 m in circumference for North and the smallest circumference was 0.65 m for the southwest. Different results were observed in the Cariri region of Paraíba, where the largest trunk diameters at nest height were facing Southwest, while the lowest values were facing West.(49)
No significant influence of nest height was observed in relation to the influence of cardinal and collateral points, although they varied between 0.86 m for the North and 2.15 m for the Southeast. Stingless bees in the Cariri region of Paraíba(49) presented a higher height of entrance holes directed to the South, while those directed to the Southwest presented nests closest to the ground.
Conclusion
Melipona subnitida is the most common stingless bee species in the Curimataú micro-region of Paraíba. Melipona asilvae, Melipona scutellaris, Partamona seridoenses, and Plebeia sp. presented the lowest frequency in the Curimataú micro-region of Paraíba. Stingless bees with occurrence in the Curimataú micro-region of Paraíba preferentially nest on Commiphora leptophloeos, with nest entrance holes directed at a variable angle between Northwest and Northeast at 1.52 m from the ground. The nesting behavior of these species is the reason to review the current model of meliponiculture adopted in this region, especially in relation to the position and height of hives in the meliponary.
Acknowledgments
The authors thank Prof. Dr. Celso Feitosa Martins (UFPB) for the taxonomic analysis and the residents of the surveyed area Antonio Paulino, Sergio, Nego Val, Dedé, Naná, Zé Luís, Sebastião Vicente, Hominho, and Nicássio for sharing their experiences with stingless bees of the region and, mainly, by their willingness to follow, alternately, the work on the trails of their properties.
References
1. Silva CI. Guia ilustrado de abelhas polinizadoras no Brasil. Universidade de São Paulo (USP). 1a Edição. São Paulo – SP, 2014.
2. Palazuelos-Ballivián JMP. (Org.). Abelhas nativas sem ferrão. São Leopoldo: Oikos; 2008. 128 p.
3. Posey D. Folk Apiculture of the Kayapó Indians of Brazil. Biotropica, [S.I.]. 1983;15(2):154–158.
4. Danforth BN. et. Al.The history of early bee diversification based on five genes plus morphology. Proceedings of the National Academy of Sciences of United States of America. 2006;103(41):15118-23.
5. Klein AM. et.al. Importance of pollinators in changing landscapes for world crops. Proceedings. Biological sciences / The Royal Society. 2007;274(1608):303-13.
6. FAO. Conservation and management of pollinators for sustainable agriculture - the international response. In: Solitary bees: conservation, rearing and management for pollination. Imprensa Universitária: Fortaleza. p.19-20;2004.
7. Allen-Wardell G. et al. The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conservation Biology. 1998; 12(1):8-17.
8. Kevan P. Pollinators as bioindicators of the state of the environment: species activity and diversity. Agriculture Ecosystems and Environment, v. 74, p. 373-393, 1999.
9. Lopes, M. Abelhas sem ferrão: a biodiversidade invisível. 2005. Disponível em: http://www.agriculturesnetwork.org/magazines/brazil/4-criacao-de-pequenosanimais/abelhas-sem-ferrao-a-biodiversidade-invisivel/at_download/article_pdf. Acesso em: 01 de Jul de 2017.
10. Foley JA. et. Al. Global consequences of land use. Science. 2005;309(5734):570-4.
11. Parmesan C. et.al. Poleward shifs in geographical ranges of butterfly species associated with regional warming. Nature. 1999;399(6736):579-583.
12. Kerr WE et al. Aspectos pouco mencionados da biodiversidade amazônica. Parcerias Estratégicas, v.12, p.20-41, 2001.
13. Kerr WE. Extinção de espécies: A grande crise biológica do momento e como afeta os meliponinios. In: V ENCONTRO SOBRE ABELHAS, 5, 1996, Ribeirão Preto, SP. Anais... Ribeirão Preto, SP, 2002, p. 4-9.
14. Castro MS. A Comunidade de abelhas (Hymenopetra, Apoidea) de uma área de caatinga arbórea entre os inselbergs de Milagres (12º53´S; 39º51´W), Bahia. 2001. (Tese Doutorado em Ecologia), Instituto de Biociências, Universidade de São Paulo. São Paulo, SP. 191p.
15. Câmara JQ. Estudos preliminares da abelha Jandaíra (Melipona subnitida D.) no município de Jandaíra – RN. Mossoró-RN: ESAM. 2004.
16. Leal IR. et al. Ecologia e conservação da Caatinga. Editora Universitária, Universidade Federal de Pernambuco, Recife, Brasil. 2003.
17. Freitas BM. et al. Diversity, threats and conservation of native bees in the Neotropics. Apidologie. 2009;40:332–346.
18. Nogueira-Neto P, Sakagami SF. Nest Structure of a Subterranean Stingless Bee – Geotrigona mombuca Smith (Meliponinae, Hymenpotera: Apoidea). An. Acad. Bras. de Ciências, [s.l.]. 1966;38(1):187-194.
19. Kerr WE. et al. Observações sobre a arquitetura dos ninhos e comportamento de algumas espécies de abelhas sem ferrão das vizinhanças de Manaus, Amazonas (Hymenoptera : Apoidea). Atas. Simp. Biota Amazônica, Manaus, AM, n. 5 (Zoologia), p. 255-309, 1967.
20. Laroca S, Almeida MC. Coexistência entre abelhas sem ferrão e formigas: Ninho de Paratrigona myrmecophila (Apidae) construído em ninho de Camponotus senex (Formicidae). Revta. Bras. Zool., Curitiba, PR.1989;6(4):671-680.
21. Imperatriz-Fonseca VL. et al. Subterranean nest structure of a stingless bee (Paratrigona subnuda Moure) (Meliponinae, Apidae, Hymenoptera). Ciência e Cultura, São Paulo, SP. 1972;24(7):662-666.
22. Camargo JMF. Biogeografia de Meliponini (Hymenoptera, Apidae, Apinae): a fauna Amazônica. In: ENCONTRO SOBRE ABELHAS, 1, 1994, Ribeirão Preto - SP, Anais... São Paulo, SP: USP, 1994, p. 46:59.
23. Nogueira-Neto P. et al. Biologia e manejo das abelhas sem ferrão. São Paulo, SP: Edição Tecnapis, 1986, 54p.
24. Vossler GG. Flower visits, nesting and nest defence behaviour of stingless bees (Apidae: Meliponini): suitability of bee species for meliponiculture in the Argentinean Chaco region. Apidologie. 2012;43:139-161.
25. Abraçado L. et al. "Magnetic material in head, thorax, and abdomen of Solenopsis substituta ants: A ferromagnetic resonance study", Journal of Magnetic Resonance. 2005;175(2):309-316.
26. Walker M. "A model for encoding of magnetic field intensity by magnetite-based magnetoreceptor cells", Journal of Theoretical Biology. 2008;250(1):85-91.
27. Begal S. et al. "Magnetic alignment in mammals and other animals", Mammalian Biology.2013;78(1):10-20.
28. Anderson JB, Vander Meer RK. Magnetic orientation in the fire ant, Solenopsis invicta. Naturwissenschaften 80, 568-570. 1993.
29. Menezes H. Própolis: uma revisão dos recentes estudos de suas propriedades farmacológicas. Arquivos do Instituto Biológico. 2005;72(3):405-411.
30. Bruening H.; Abelha Jandaíra. 3a ed. Natal: SEBRAE/ RN, 2006.
31. Serra BD. V. Abundância, distribuição espacial de ninhos de abelhas Meliponina (Hymenoptera, Apidae, Apini) e espécies vegetais utilizadas para nidificação em áreas de cerrado do Maranhão. Iheringia, Série Zoologia. 2009;99(1):12-17.
32. Kerr W. E. et al. Abelha Uruçu : Biologia, Manejo e Conservação – Belo Horizonte-MG: Acangaú, 1996. 144 p.: il., (Coleção Manejo da vida silvestre; 2).
33. Rosso L. J. M. et al. Meliponicultura em Brasil I: situacion em 2001 y perspectivas. In: Seminário Mexicano sobre Abejas sin Aguijón, 2., Yucatán, 2001. Anais... Yucatán, 2001. p. 28-35.
34. Campos LAO, Peruquetti RC. Biologia e criação de abelhas sem ferrão. Informe Técnico. Viçosa: UFV. 1999; n. 82, 38p.
35. Camargo JMF, Pedro SRM. Meliponini Lepeletier, 1836. In Moure, J. S., Urban, D., Melo, G. A. R. (Orgs). 2013. Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - online version. Disponível em: http://www.moure.cria.org.br/catalogue. Acessado em 05/07/2017
36. Waldschmidt AM. Meliponicultura na Bahia. In: Congresso Baiano de Apicultura, 2. 2002, Paulo Afonso, BA. Anais... Paulo Afonso: p. 166-168. 2002.
37. Sakagami SF. Stingless bees. In: Herman, H. R. ed. Social Insects III. London, Academic Press, p. 361-423, 1982.
38. AQUINO IS. Abelhas Nativas da Paraíba. 1ª edição. João Pessoa. Editora Universitária - UFPB. 2006. 91p.
39. Lorenzon MCA. et al. Flora visitada pelas abelhas eussociais (Hymenoptera, Apidae) na Serra da Capivara, em caatinga do Sul do Piauí. Neotropical Entomology. 2003;32(1):27-36.
40. Martins CF.et al. Espécies arbóreas utilizadas para nidificação por abelhas sem ferrão na caatinga (Seridó, PB; João Câmara, RN). Biota Neotropica. 2004;4(2):1-8.
41. Antonini Y. Efeitos de variáveis ecológicas na ocorrência de Melipona quadrifasciata (Apidae, Meliponini) em fragmentos urbanos e rurais. 2002. Tese de doutorado, Universidade Federal de Minas Gerais, Belo Horizonte.
42. Ferreira R.LC, Vale AB. Subsídios básicos para o manejo florestal da caatinga. Anais do 2º Congresso Nacional sobre Essências Nativas:368-375. 1992.
43. Villas-Bôas J. Manual Tecnológico: Mel de Abelhas sem Ferrão. Brasília – DF. Instituto Sociedade, População e Natureza (ISPN). Brasil, 2012.
44. Werneck HA.; Faria-Mucci GM. Abelhas sem Ferrão (Hymenoptera:Apidae, Meliponini) da Estação Ecológica de Água Limpa, Cataguases-MG, Brasil. EntomoBrasilis. 2014;7(2):164-166.
45. Pedro SRM.; Camargo JM. F. Meliponini Neotropicais: o gênero Partamona Schwarz, 1939 (Hymenoptera, Apidae). Revista Brasileira de Entomologia, 47(supl. 1): 1-117. 2003.
46. Pioker-Hara FC. Determinantes da densidade e distribuição de ninhos e diversidade de espécies de meliponineos (Apidae, Meliponini) em áreas de cerrado de Itirapina, SP. Tese de doutorado em Ecologia. Universidade de São Paulo, USP, Brasil. 2011.
47. Lacerda DC. O. Influência da Orientação Magnética na Nidificação de Abelhas Nativas na Zona da Mata Pernambucana. 2017. Dissertação (Mestrado em Ciências Agrarias "Agroecologia"). Universidade Federal da Paraíba-UFPB, Bananeiras-PB, 77p.
48. Souza SGX. et. al. As abelhas sem ferrão (Apidae; Meliponina) residentes no campus Federação/Ondina da Universidade Federal da Bahia, Salvador. Candombá - Revista Virtual. 2005;1(1):57–69.
49. Silva FJA. Preferência de Nidificação em Abelhas Indígenas. 2016. Dissertação (Mestrado em Ciências Agrarias "Agroecologia"). Universidade Federal da Paraíba-UFPB, Bananeiras-PB, 90p.