
Resumo
As melíponas, além de excelentes produtoras de mel, prestam um grande serviço à humanidade através da sua capacidade de polinizar plantas e sua consequente preservação de ecossistemas. Nessa relação interespecífica harmônica, as abelhas são beneficiadas pelas espécies vegetais pelo provimento do alimento (néctar e pólen), além de utilizarem seus galhos e troncos para se alojarem. Porém, a ação antrópica predatória tem colocado em risco a sobrevivência das melíponas, causando danos ambientais irreversíveis e comprometendo a preservação dos recursos naturais. Este trabalho objetivou identificar os hábitos de nidificação das abelhas nativas em condições naturais. Por meio de trilhas aleatórias na microrregião do Curimataú paraibano foi possível identificar os pontos de nidificação das espécies de meliponíneos em seu habitat natural. Foram identificados 60 ninhos de melíponas nidificando em 12 espécies vegetais diferentes, sendo a Commiphora leptophloeos a hospedeira de maior preferência (46,66%), com 55% dos orifícios de entrada apontados para orientações magnéticas entre o Nordeste e Noroeste; com maior escolha por hospedeiras (árvores) [85%]; apresentando preferência por nidificação em vegetais hospedeiros com circunferência de 0,98 m de diâmetro, orifício de entrada dos ninhos com 7,77 mm de diâmetro e uma altura do solo de 1,52 m; Os dados obtidos revelam pouca variedade de espécies melíponas na região Curimataú paraibano, com destaque para Melipona subnitida. As espécies Melipona asilvae, Melipona scutellaris, Partomona seridoenses e Plebeia sp. devem ter uma atenção especial para não entrarem em extinção nessa região.
Palavras-chave: Curimataú. Habitat natural. Melíponas. Nidificação. Preferência.
Abstract
Native bees, besides excellent producers of honey, lend a great legacy to humanity through their ability to pollinate plants and the consequent preservation of ecosystems. In this interspecific relation, bees are benefited by plant species by the provision of food (nectar and pollen), besides using their branches and trunks for lodging. However, the anthropogenic action has jeopardized the survival of stingless bees, causing irreversible environmental damage in the preservation of natural resources. This study aimed to identify the nesting habits of native bees under natural conditions. Random trails in the Curimataú micro-region of Paraíba allowed identifying nesting sites of indigenous species in their natural habitat. Sixty honeycomb nests were identified in 12 different plant species, being Commiphora leptophloeos the most preferred host (46.66%), with 55% of the entrance holes pointed to magnetic orientations between the Northeast and Northwest; a greater choice by host trees (85%); preference for nesting in host plants with a circumference of 0.98 m in diameter, entrance hole of nests of 7.77 mm in diameter, and height from the ground of 1.52 m. The obtained data showed little variety of stingless bee species in the Curimataú region of Paraíba, especially Melipona subnitida. The species Melipona asilvae, Melipona scutellaris, Partamona seridoenses, and Plebeia sp. should receive special attention not to become extinct in this region.
Keywords: Curimataú. Natural habitat. Nesting. Preference. Stingless bees.
Seção: Zootecnia
Recebido
25 de maio de 2019.
Aceito
18 de junho de 2019.
Publicado
6 de agosto de 2020.
www.revistas.ufg.br/vet
Como citar - disponível no site, na página do artigo.
Introdução
Os meliponíneos, conhecidos também como abelhas melíponas, sem ferrão ou indígenas, habitam o Planeta Terra há cerca de 125 milhões de anos(1). A meliponicultura é uma atividade bastante antiga praticada desde os primórdios das civilizações no Antigo Egito(2). No Brasil, a criação de abelhas melíponas foi iniciada pelos índios(3) antes mesmo do descobrimento e da introdução da Apis mellifera L. (Hymenoptera: Apidae)
As abelhas prestam um grande serviço à humanidade através da sua capacidade de polinização, pois são insetos especializados na polinização das plantas(4), colaborando para a preservação dos ecossistemas e gerando um acréscimo na produção de espécies vegetais comerciais(5). Além disso, são os principais agentes polinizadores do planeta. Estima-se que, aproximadamente, 80% das espécies vegetais cultivadas no mundo sejam polinizadas por alguma espécie de abelha, 19% por moscas, 6,5% por morcegos, 5% por vespas, 5% por besouros, 4% por pássaros e 4% por borboletas e mariposas(6).
O uso indiscriminado de agrotóxicos tem colocado em risco a fauna e a flora através dos seus efeitos nocivos aos insetos polinizadores dos agroecossistemas(7). Uma das maiores consequências dessa prática é a direta diminuição da população de abelhas, além dos efeitos (indiretos) de perdas econômicas decorrentes do declínio desses polinizadores(8).
A ação antrópica, ou seja, a ação predatória do homem ao longo de décadas tem causado danos ambientais irreversíveis na preservação dos recursos naturais. Os avanços das fronteiras agrícolas elevaram os índices de desmatamento e uso indiscriminado de agrotóxicos, dificultando a fixação das abelhas nativas em seus habitats. Com isso, os meliponíneos encontram-se em processo acelerado de desaparecimento, provocado principalmente pela perda de qualidade ambiental causada pela derrubada de florestas nativas(9). Observa-se, assim, que a degradação e destruição do meio ambiente parece ser o principal fator do declínio da biodiversidade(10). Outros fatores, entretanto, vêm causando preocupação na preservação destas espécies, como o aquecimento global(11).
Uma ameaça iminente e comprometedora ao futuro da meliponicultura é atribuída aos "meleiros", pessoas que exploram irregularmente o comércio do mel de abelhas indígenas. Essas pessoas derrubam as árvores a procura de abelhas nativas, retiram o mel de forma rudimentar, destroem os discos de crias das colmeias e os que sobram jogam no solo, sendo devorados pelas formigas. Isso provoca rapidamente a dizimação das melíponas(12, 13) pela destruição dos seus ninhos por meio dessa ação indiscriminada e predatória praticadas pelos meleiros(14).
A criação das abelhas indígenas, muitas vezes realizada pelos produtores de forma empírica, gera uma demanda de pesquisas que contribuam para uma produção com técnicas de manejo(15) adequadas. O presente trabalho teve como objetivo identificar os hábitos de nidificação de meliponíneos em condições naturais de campo, quanto a sua ocorrência, localização, substratos vegetais, características dos substratos, arquitetura e preferências de nidificação, levando em consideração ainda a orientação magnética na microrregião do Curimataú paraibano, além de propor especificações técnicas que venham a ser utilizadas na confecção de caixas padrão, na orientação do posicionamento das mesmas em relação aos pontos cardeais e no manejo racional e humanizado das melíponas de forma sustentável.
Os meliponíneos são abelhas dóceis que vivem em colônias e são de fácil manejo(17). A compreensão do processo de nidificação das meliponinas é de fundamental importância para a preservação das espécies e para um planejamento minucioso de um sistema de produção sustentável, levando em consideração a região de ocorrências das espécies e as características do habitat em que estão inseridas. Com base nessa compreensão, vários autores(18-22) descreveram a preferência de diversos hábitos de nidificação para as espécies de meliponinas.
O diâmetro do tronco de árvores que abrigam os ninhos das melíponas dá uma ideia do diâmetro da cavidade preexistente ocupada pelo ninho e pode variar de espécie para espécie. A maioria das espécies depende dessas cavidades preexistentes, geralmente em ocos de árvores vivas ou mortas para construírem seus ninhos(23), e o seu diâmetro interno sofre variação de acordo com a espécie nidificada(24).
A capacidade de orientação dos seres vivos é um fator determinante na sobrevivência das espécies, seja no deslocamento, na coleta de alimentos ou na reprodução. Assim, as mais variadas espécies desenvolvem mecanismos de adaptação ao meio. Dentre tais habilidades, merece destaque a teoria da capacidade de detectar o campo magnético da Terra para orientação e navegação, comumente aceita por vários pesquisadores(25-27), muito embora pouco conhecida em animais(28).
O substrato é mais um parâmetro que sofre variações quanto aos hábitos de nidificação das melíponas. Algumas vezes dentro do mesmo gênero de Meliponina são encontrados vários hábitos de nidificação(22). Não são conhecidos os fatores que direcionam a preferência das abelhas por uma determinada fonte vegetal(29), porém relatos demonstram que elas são seletivas nessa escolha. De forma geral, as melíponas dependem das árvores para moradia e alimentação(30, 24), construindo seus ninhos em cavidades preexistentes. A região de ocorrência de todas as espécies de meliponíneos deve ser respeitada dentro de uma exploração racional, evitando, assim, a degradação daquele ecossistema. A compreensão do padrão de sua distribuição e abundância é considerada importante(31) para que se desenvolvam estratégias de conservação, a fim de reduzir os efeitos da fragmentação de habitats sobre a estrutura genética e ecológica de populações de meliponíneos.
As abelhas fazem parte da ordem Hymenoptera e as melíponas estão reunidas na superfamília Apoidea sendo a mesma subdividida em oito famílias: Andrenidae, Anthophoridae, Colletidae, Halictidae, Melittidae, Megachilidae, Oxaeidae e Apidae. A família Apidae por sua vez se divide em quatro subfamílias: Bombinae, Euglossinae, Apinae e Meliponinae. A subfamília Meliponinae encontra-se dividida em duas tribos: Meliponini e Trigonini. As abelhas de ambas as tribos podem ser chamadas de meliponíneos(32). Dentro da subfamília Meliponinae, há 52 gêneros com um total de 400 espécies(33) espalhadas em todo o mundo, desde o estado do Rio Grande do Sul no Brasil até o centro do México, África, Índia, Malásia, Indonésia e Austrália(32).
O Brasil é um dos principais locais de ocorrência dos meliponíneos, onde eles exercem grande influência na manutenção dos ecossistemas. Até o momento foram descritos 33 gêneros de meliponíneos Neotropicais no País, compreendendo 300 espécies(34), podendo este número variar entre 350 e 400 espécies(35). Porém, acredita-se que a quantidade de espécies seja bem maior, com elevada diversidade de formas, tamanho e hábitos de nidificação(36). Pelo menos 100 espécies meliponíneos estão em perigo de extinção devido à destruição de seus habitats pelo homem(32).
A tribo Meliponini é exclusiva das Américas, na região Neotropical (do sul da América do Sul até as montanhas do norte do México, na América Central), com aproximadamente 26 espécies e numerosas subespécies e está representada por um único gênero: Melipona. No Brasil, estão presentes mais ou menos 20 espécies: Melipona scutellaris (uruçu), M. mandacaia (a mandaçaia do Nordeste do Brasil), M. asilvai (manduri), M. quinquefasciata (uruçu do chão), M. subnitida (jandaíra), entre outras(37). Na Paraíba, a ocorrência da tribo Meliponini está distribuída nas microrregiões do Litoral, Curimataú, Borborema e Sertão com cinco espécies: Melipona subnitida, M. quadrifasciata, M. asilvae, M. compressipes fasciculada e M. scutellaris(38).
A tribo Trigonini constitui um grupo muito diversificado de abelhas sem ferrão, abrangendo o maior número de gêneros existentes na região Neotropical, sendo possível encontrar diversos gêneros: Lestrimelitta, Hypotrigona, Cephalotrigona, Oxytrigona, Schwarziana, Scaura, Plebeia, Partamona, Paratrigona, Scaptotrigona, Nannotrigona, Tetragonisca, Trigona, Geotrigona, Frieseomelitta, e inúmeras espécies; em torno de 120(37). Na Paraíba foram encontradas 13 espécies da tribo Trigonini, distribuídas em oito gêneros diferentes: Frieseomellita, Trigona, Scaptotrigrona, Partomona, Tetragonisca, Leurotrigona, Olebeia e Nannotrigona(38).
A Caatinga é um bioma único, localizado em um clima semiárido, e é o principal ecossistema existente na região Nordeste do Brasil, berço natural das melíponas. Esta região apresenta uma grande variedade de paisagens com relativa riqueza biológica e endêmica, regida por secas estacionais e periódicas que se renova a cada ciclo de chuvas. Esse bioma vem passando por uma extensa devastação ambiental provocado por um processo antrópico do uso indiscriminado dos seus recursos naturais(16).
O Curimataú paraibano constitui-se em uma microrregião dentro do bioma Caatinga que é conhecida pela presença de abelhas indígenas. Municípios dessa localidade, como Araruna, Cacimba de Dentro, Damião e Cuité, possuem uma vasta área predominantemente de Caatinga e clima semiárido, marcado por baixos índices pluviométricos, curto período de chuva, mal distribuída, com fauna e flora ameaçadas pela ação antrópica. Portanto, conhecer os hábitos de nidificação de abelhas melíponas nessa região, torna-se imprescindível para que os meliponicultores possam aprimorar suas técnicas de manejo. Este trabalho objetivou identificar os hábitos de nidificação de abelhas nativas em condições naturais na microrregião Curimataú paraibano.
Material e métodos
Este trabalho foi realizado na região do Curimataú paraibano. Para efeito de amostragem os dados foram coletados em quatro municípios: Araruna (06°33'30"S – 35°44'30"W), Damião (06°37'54"S – 35°54'18"W), Cuité (06°29'01"S – 36°09'13"W) e Cacimba de Dentro (06°38'21"S – 35°46'42"W) totalizando uma área de 1.336,9 km², durante o período de setembro a novembro de 2016. Os locais de nidificações foram previamente identificados através de entrevistas com agricultores familiares habitantes nas áreas de coleta.
Através de trilhas aleatórias, iniciou-se, com o auxílio de informantes nativos da região, o levantamento dos possíveis locais de nidificação das abelhas nativas do Curimataú paraibano. A identificação botânica das árvores hospedeiras foi realizada pelos colaboradores por meio do nome vulgar das espécies e confirmadas por meio de uma busca em sites especializados da internet (Tópicos e Flora do Brasil).
Os locais de nidificação foram catalogados utilizando-se os sistemas do Sistema de Navegação Global por Satélite (GPS) por meio de um aparelho Garmin® etrex 30, possibilitando identificar as coordenadas geográficas de cada local de nidificação e a orientação magnética (pontos cardeais e colaterais) da entrada do orifício dos ninhos preferidos pelas abelhas nativas. Outros parâmetros foram avaliados para uma melhor caracterização e identificação da preferência das abelhas nativas no ato da escolha dos locais de nidificação. Utilizou-se a metodologia descrita por Serra(31), medindo-se o diâmetro do orifício de entrada do ninho com um paquímetro Lotus®, o diâmetro da árvore no local do orifício de entrada com uma fita métrica Stanley® e a altura do orifício de entrada do ninho em relação à altura do solo com auxílio de trena Irwin® de 5 metros. Para a captura de amostras com altura de orifícios superior a 2 metros foi utilizada uma rede entomológica.
Foi coletada uma amostra com cinco espécimes de abelhas nativas em cada local de nidificação acondicionando-as em um recipiente de acrílico (50 ml), com tampa, contendo 30 ml de álcool (70%). Os recipientes foram devidamente identificados, e enviados ao Laboratório de Entomologia (LE) do Departamento de Sistemática e Ecologia (DES) da Universidade Federal da Paraíba (UFPB) para serem submetidas à análise taxonômica.
O Qui-quadrado foi utilizado para as frequências de ocorrências das espécies, enquanto, que o Duncan, após a Anova, está sendo utilizado para comparar as médias de diâmetro do orifício de entrada, diâmetro do tronco e altura do ninho em função das espécies e da orientação.
Resultados e discussão
Foram encontrados 60 ninhos de abelhas melíponas em quatro municípios diferentes do Curimataú paraibano (Araruna, Cacimba de Dentro, Damião e Cuité). As abelhas encontradas (Tribos Meliponini e Trigonini) corresponderam a oito espécies diferentes: Melipona subnitida Ducke, 1910, Melipona asilvai Moure, 1971 (Hymenoptera: Apidae), Melipona scutellaris Latreille, 1811 (Hymenopetra: Apidae), Frieseomelitta doederleini Friese, 1900 (Hymenopetra: Apidae), Frieseomelitta francoi Moure, 1946 (Hymenopetra: Apidae), Partamona seridoenses Pedro & Camargo, 2003 (Hymenopetra: Apidae), Plebeia sp. e Scaptotrigona aff. depilis Moure, 1942 (Hymenoptera, Apidae); sendo a M. subnitida a mais frequente (36,67%), seguida pela F. doederleini (25%), a Frieseomelitta francoi Moure, 1946 (Hymenoptera, Apidae) e a S. aff. depilis, ambas com 11,67% (Tabela 1). Em estudo realizado na Serra da Capivara-PI,(39) também foi observada a ocorrência dessas espécies.
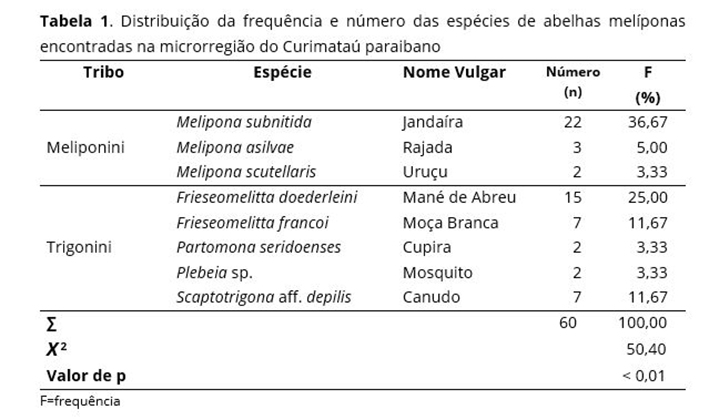
Das 18 espécies de abelhas nativas que habitam na Paraíba(38), oito delas encontraram-se distribuídas na área de coleta (microrregião Curimataú paraibano), tendo como espécie predominante de maior ocorrência na área estudada Melipona subnitida com 36,67% dos ninhos encontrados. Isso confirma a predominância dessa espécie na microrregião Curimataú(38), seguida pela Frieseomelitta doederleini (25%) e Scaptotrigona aff. postica Latreille, 1807 (Hymenopetra: Apidae) (11,67%). As abelhas Melipona scutellaris, Partamona seridoenses e P. sp são as de menor ocorrência (todas com 3,33%) [Tabela 1]. Considerando que as espécies de meliponíneos encontram-se em processo acelerado de desaparecimento(9) provocado, principalmente, pela perda de habitat pelo desmatamento de florestas nativas, ambiente natural dessas espécies, a sua baixa incidência sugere que se faça um programa de educação ambiental junto às comunidades locais, particularmente junto aos meliponicultores. Em pesquisa de campo realizada no município de João Câmara, Rio Grande do Norte, Martins et al.(40) encontraram sete espécies de abelhas melíponas, tendo a M. subnitida com maior ocorrência (57,3%), seguida pela Melipona asilvai Moure, 1971 (Hymenoptera: Apidae) (17,2%) e a Frieseomelitta dispar Moure, 1950 (Hymenoptera: Apidae) (13,7%).
Diante de centenas de espécies de árvores existentes na região Curimataú, observa-se que as melíponas utilizam um número pequeno para nidificação; neste estudo, apenas 12 espécies (Tabela 2). Espécies de abelhas sem ferrão são seletivas e optam por árvores específicas para nidificar(41).
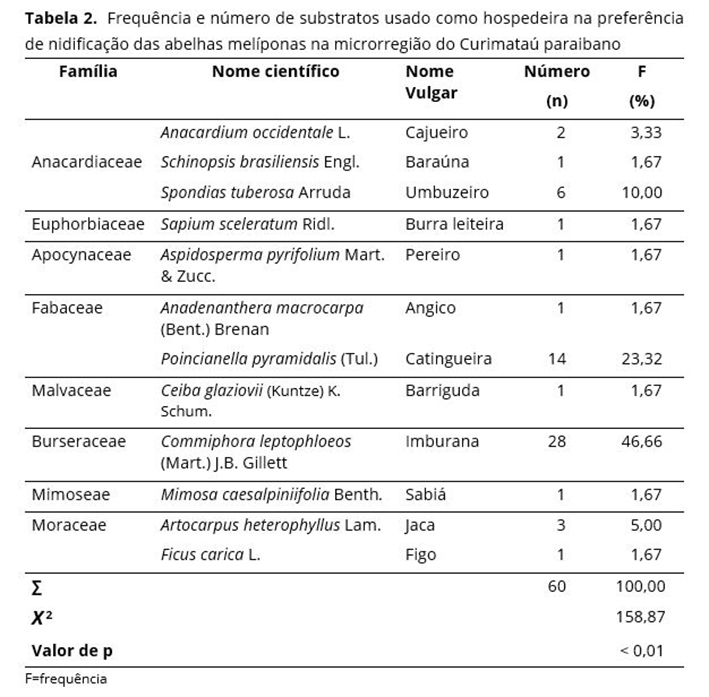
Este estudo mostra que abelhas sem ferrão nidificam em diferentes espécies arbóreas. Entretanto, 69,98% dos ninhos foram observados em apenas duas espécies de árvores: Commiphora leptophloeos (Mart.) J.B. Gillett, com 46,66%; seguida pela Poincianella pyramidalis (Tul.), com 23,32% (Tabela 2). Dados semelhantes foram encontrados em estudos fitosociológicos realizados em uma área de Caatinga de crescimento secundário, na região do Seridó(42), onde Poincianella pyramidalis apresentou as maiores densidades e frequências. A espécie C. leptophoeos é conhecida por apresentar o seu tronco com cavidade oca, condição essencial para instalação dos ninhos das melíponas. Resultados semelhantes na preferência das espécies vegetais hospedeiras com abelhas sem ferrão na Caatinga (Seridó, PB; João Câmara, RN) foram observados(40) para C. leptophoeos e Poincianella pyramidalis com frequência de 75,0%, bem próxima à encontrada neste trabalho (69,98%).
O conhecimento da preferência das abelhas melíponas nas escolhas dos locais para nidificarem vão além de uma cavidade e uma boa florada. Deve-se levar em consideração, também, a orientação magnética, ou seja, para onde vai estar voltado o orifício de entrada do ninho.
Observa-se que há certa preferência na orientação magnética das melíponas encontradas na microrregião do Curimataú paraibano. Os pontos cardeais e colaterais com orientação variável entre Nordeste e Noroeste (Tabela 3) apresentam maior escolha, com uma frequência de 55%.
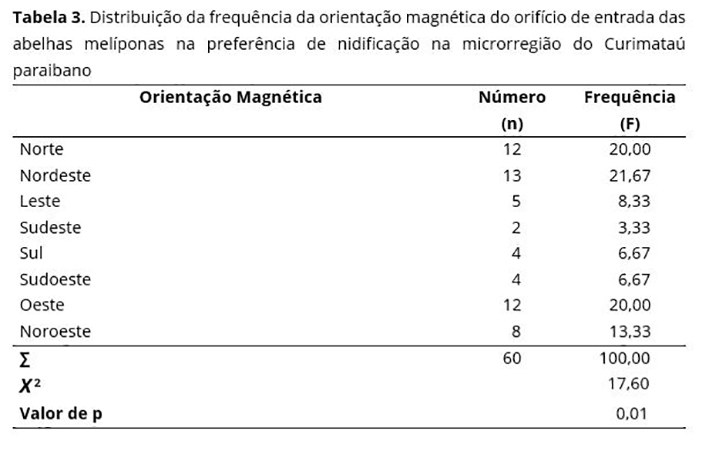
Os menores resultados para orientação magnética foram observados para as posições Sudeste (SE) 3,33%, Sul (S) 6,67% e Sudoeste (SO) 6,67% somando um total de 16,67% .
Em seu habitat natural, as abelhas sem ferrão buscam a instalação de seus ninhos em locais camuflados. Na sua maioria, essas abelhas usam oco de árvores vivas, árvores mortas, tronco seco, pedras, cupim, formigueiros, paredes telhas etc. Porém, é predominante a nidificação em troncos de árvores vivas(43).
Este trabalho revela que 85% das melíponas nidificam em cavidade de árvores vivas, seguida por árvore morta (6,67%), tronco seco (5,0%) e cupinzeiro (3,33%) (Tabela 4). Em estudo município de Cataguases-MG, 20 dos 21 ninhos foram encontrados em substrato arbóreo, apenas um ninho de T. angustula foi construído no barranco(44). Ficando evidente a preferências das melíponas por árvores vivas para nidificarem.
Apenas 3,33% das espécies encontradas nidificaram em cupinzeiros (Tabela 4). Esses cupinzeiros, por sua vez, estavam ocupados por P. seridoenses, abelha que apresenta hábito específico para nidificação em cupinzeiros(45).

As melíponas apresentam características próprias que podem sofrer influência no momento da nidificação em relação ao diâmetro do orifício de entrada, diâmetro do tronco e altura no ninho. Observa-se que o tamanho do orifício dos ninhos das melíponas estudadas diferiu entre espécies (Tabela 5); já em relação à altura do ninho, não houve diferença entre espécies.
O diâmetro do orifício de entrada dos ninhos das melíponas pode sofrer variação quando levado em consideração o porte de cada espécie; ou seja, quanto maior o porte, maior o orifício de entrada. Observa-se a ocorrência de uma diferença significativa do diâmetro do orifício de entrada quando comparada com as espécies estudadas (Tabela 5) em que o maior diâmetro foi da Scaptotrigona com 10,43 mm, não diferindo estatisticamente da M. scutellaris (9,0 mm), M. subnitida (8,45 mm), e P. seridoensis (7,50 mm).
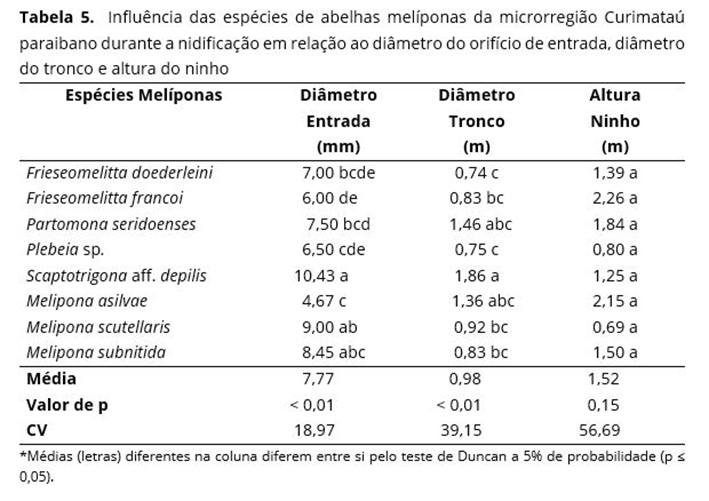
Resultados intermediários para diâmetro do orifício de entrada foram registrados pela P. seridoensis com 7,5 mm, F. doederleini com 7 mm e F. francoi com 6 mm; o menor resultado foi da M. asilvae, (4,67 mm) (Tabela 5). Os meliponíneos constroem seus ninhos tipicamente em cavidades preexistente de árvores nativas, sendo que o diâmetro interno varia de acordo com a espécie nidificada(24).
O diâmetro do tronco sofreu influência das espécies de melíponas estudadas em que os maiores resultados foram atribuídos a S. aff. depilis (1,86 m), P. seridoenses (1,46 m), M. asilvae (1,36 m) e M. scutellaris (0,92 m), diferindo M. subnitida, com variação de 0,74 a 0,83 m (Tabela 5); resultados diferentes (dos encontrados neste trabalho) mostram que os diâmetros encontrados ficaram entre 0,39 a 0,50 m(40).
O diâmetro do tronco pode estar diretamente relacionado ao volume de ocupação utilizado pelas melíponas, que pode variar quanto à espécie em relação as suas exigências para construir seu ninho. A capacidade da cavidade preexistente pode ser determinante na escolha do local de nidificação(46). O volume da colmeia é um fator determinante, podendo sofrer variações de acordo com a região(32). Geralmente, uma colmeia de M. subnitida requer um volume de 15 litros.
A altura do ninho não sofreu variação significativa quando comparada em relação às espécies de melíponas, embora tenham apresentado uma variação de 0,80 m para Plebeia sp. e 2,6 m para F. francoi (Tabela 5), fato que provocou um alto coeficiente de variação. Houve uma influência significativa das espécies vegetais hospedeiras da microrregião Curimataú paraibano em relação ao diâmetro do orifício de entrada dos ninhos e diâmetro do tronco dos ninhos encontrados em seu habitat natural (Tabela 6). É possível constatar também que não ocorre diferença significativa em relação à altura do ninho.
Os maiores orifícios de entrada foram encontrados nas espécies A. heterophyllus, F. carica, ambas com 11 mm; A. occidentale com 10 mm, C. glaziovii e S. sceleratum, ambas com 9 mm. Os menores resultados encontrados ficaram com A. macrocarpa, A. pyrifolium e S. brasiliensis, com 4, 5 e 4 mm, respectivamente (Tabela 6).
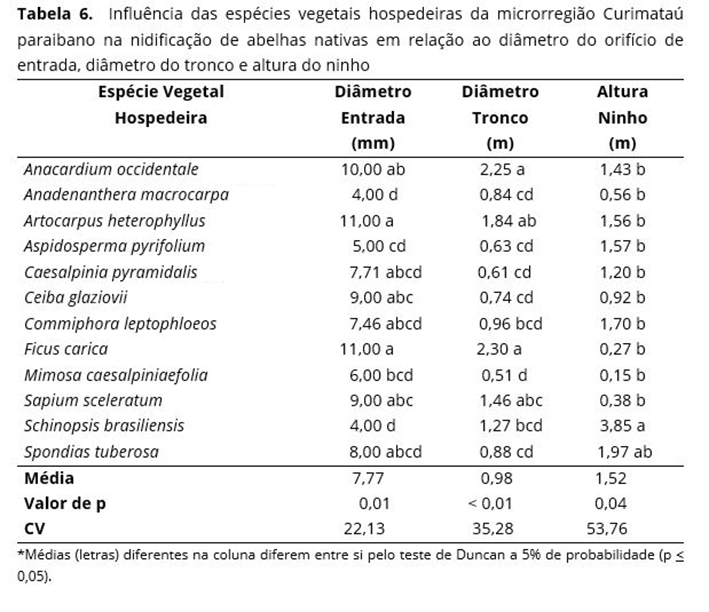
Ao comparar o diâmetro do tronco das árvores em relação à influência dos vegetais hospedeiros (Tabela 6), observa-se que os maiores diâmetros foram apresentados pela A. occidentale e F. carica com 2,25 m e 2,30 m, respectivamente, diferindo estatisticamente da M. caesalpiniaefolia com 0,51 m o menor resultado. Em um estudo realizado na Zona da Mata pernambucana, 50% das abelhas (meliponíneos) nidificam em árvores com diâmetro de inferior a 50%; 30% com diâmetro entre 1 a 1,5 m, e 20% com diâmetro entre 2 a 2,5 m(47). A variação no diâmetro das árvores escolhidas para nidificação das espécies de melíponas encontradas no Curimataú paraibano deve estar relacionada ao tamanho das árvores presentes na área de estudo, com características variadas quanto ao porte e arquitetura dessas plantas.
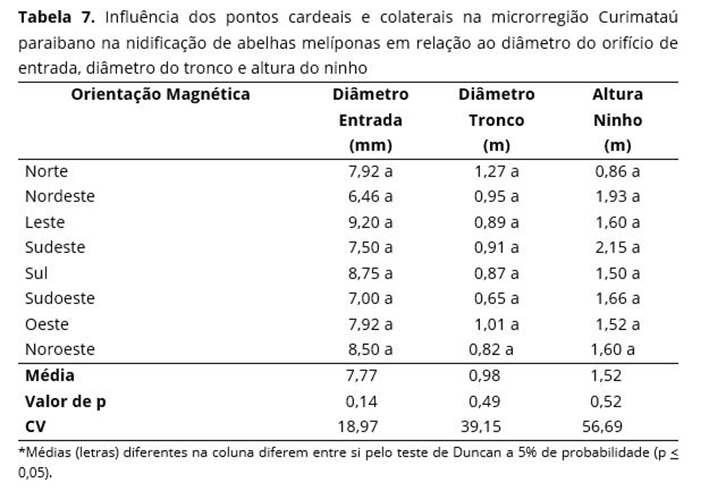
A altura dos ninhos encontrados não sofreram diferenças significativas quanto às espécies de árvores hospedeiras, mesmo apresentando uma amplitude máxima de 3,85 m para S. brasiliensis e mínima para M. caesalpiniaefolia, com 0,15 m de altura do ninho em relação com o solo. Resultados semelhantes foram encontrados em um levantamento realizado na Bahia com melíponas(48), onde a altura dos ninhos variou entre 0,5 a 3,8 m. É provável que a altura dos ninhos das melíponas não esteja relacionada com o porte ou o tipo de árvores, mas sim, com as características de nidificação de cada espécie de abelha. A altura do orifício de entrada dos ninhos em espécies vegetais hospedeiras nas posições mais alta ou mais baixa é realizada de forma casual(31). É possível constatar que os pontos cardeais e colaterais não influenciam o diâmetro do orifício de entrada, diâmetro do tronco e altura do ninho (Tabela 7).
Apesar de não diferir estatisticamente, o diâmetro do orifício de entrada apresentou os maiores resultados para os pontos que se encontravam voltados para o Leste, Sul e Noroeste com 9,2, 8,75 e 8,5 mm, respectivamente; seguido dos menores resultados para os pontos Nordeste, Sudeste e Sudoeste 6,46; 7,5 e 7,0 mm, respectivamente (Tabela 7).
Observa-se que o diâmetro do tronco das árvores hospedeiras não sofreu influência significativa quando comparada com os pontos cardeais e colaterais (Tabela 7), embora se verifique que o maior diâmetro foi de 1,27 m de circunferência para a direção Norte (N) e a menor circunferência foi de 0,65 m para o Sudoeste (SO). Resultados diferentes foram observados no Cariri paraibano, onde os maiores diâmetros de tronco na altura dos ninhos estavam voltados para o Sudoeste (SO) e os menores valores estavam voltados para o Oeste [O](49).
Não houve influência significativa da altura dos ninhos encontrados em relação à influência dos pontos cardeais e colaterais, embora tenha sofrido uma variação entre 0,86 m para o Norte e 2,15 m para o Sudeste. As abelhas sem ferrão no Cariri paraibano(49) com alvados apontados para o Sul (S) apresentam seus ninhos com maior altura (em relação ao solo); já os apontados para o Sudoeste (SO), apresentam os ninhos mais próximos ao solo.
Conclusão
Melipona subnitida é a espécie melípona de maior ocorrência na microrregião do Curimataú paraibano. Melipona asilvae, Melipona scutellaris, Partomona seridoenses e Plebeia sp. são as espécies de menor frequência na microrregião do Curimataú paraibano. As melíponas com ocorrência na microrregião do Curimataú paraibano nidificam preferencialmente em Commiphora leptophoeos, com orifícios de entrada dos ninhos direcionados para uma angulação variável entre Noroeste e Nordeste, a 1,52 m do solo. Esse comportamento de nidificação dessas espécies é motivo para rever-se o atual modelo de meliponicultura adotado nessa região, especialmente em relação à posição e altura das colmeias no meliponário.
Agradecimentos
Os autores agradecem ao Prof. Dr. Celso Feitosa Martins (UFPB) pela análise taxonômica e aos moradores da área pesquisada, Antônio Paulino, Sérgio, Nego Val, Dedé, Naná, Zé Luís, Sebastião Vicente, Hominho e Nicássio, pelo compartilhamento de suas vivências com as abelhas melíponas da região e, principalmente, pela disposição em acompanhar – alternadamente – os trabalhos nas trilhas de suas propriedades.
Referências
1. Silva CI. Guia ilustrado de abelhas polinizadoras no Brasil. Universidade de São Paulo (USP). 1a Edição. São Paulo – SP, 2014.
2. Palazuelos-Ballivián JMP. (Org.). Abelhas nativas sem ferrão. São Leopoldo: Oikos; 2008. 128 p.
3. Posey D. Folk Apiculture of the Kayapó Indians of Brazil. Biotropica, [S.I.]. 1983;15(2):154–158.
4. Danforth BN. et. Al.The history of early bee diversification based on five genes plus morphology. Proceedings of the National Academy of Sciences of United States of America. 2006;103(41):15118-23.
5. Klein AM. et.al. Importance of pollinators in changing landscapes for world crops. Proceedings. Biological sciences / The Royal Society. 2007;274(1608):303-13.
6. FAO. Conservation and management of pollinators for sustainable agriculture - the international response. In: Solitary bees: conservation, rearing and management for pollination. Imprensa Universitária: Fortaleza. p.19-20;2004.
7. Allen-Wardell G. et al. The potential consequences of pollinator declines on the conservation of biodiversity and stability of food crop yields. Conservation Biology. 1998; 12(1):8-17.
8. Kevan P. Pollinators as bioindicators of the state of the environment: species activity and diversity. Agriculture Ecosystems and Environment, v. 74, p. 373-393, 1999.
9. Lopes, M. Abelhas sem ferrão: a biodiversidade invisível. 2005. Disponível em: http://www.agriculturesnetwork.org/magazines/brazil/4-criacao-de-pequenosanimais/abelhas-sem-ferrao-a-biodiversidade-invisivel/at_download/article_pdf. Acesso em: 01 de Jul de 2017.
10. Foley JA. et. Al. Global consequences of land use. Science. 2005;309(5734):570-4.
11. Parmesan C. et.al. Poleward shifs in geographical ranges of butterfly species associated with regional warming. Nature. 1999;399(6736):579-583.
12. Kerr WE et al. Aspectos pouco mencionados da biodiversidade amazônica. Parcerias Estratégicas, v.12, p.20-41, 2001.
13. Kerr WE. Extinção de espécies: A grande crise biológica do momento e como afeta os meliponinios. In: V ENCONTRO SOBRE ABELHAS, 5, 1996, Ribeirão Preto, SP. Anais... Ribeirão Preto, SP, 2002, p. 4-9.
14. Castro MS. A Comunidade de abelhas (Hymenopetra, Apoidea) de uma área de caatinga arbórea entre os inselbergs de Milagres (12º53´S; 39º51´W), Bahia. 2001. (Tese Doutorado em Ecologia), Instituto de Biociências, Universidade de São Paulo. São Paulo, SP. 191p.
15. Câmara JQ. Estudos preliminares da abelha Jandaíra (Melipona subnitida D.) no município de Jandaíra – RN. Mossoró-RN: ESAM. 2004.
16. Leal IR. et al. Ecologia e conservação da Caatinga. Editora Universitária, Universidade Federal de Pernambuco, Recife, Brasil. 2003.
17. Freitas BM. et al. Diversity, threats and conservation of native bees in the Neotropics. Apidologie. 2009;40:332–346.
18. Nogueira-Neto P, Sakagami SF. Nest Structure of a Subterranean Stingless Bee – Geotrigona mombuca Smith (Meliponinae, Hymenpotera: Apoidea). An. Acad. Bras. de Ciências, [s.l.]. 1966;38(1):187-194.
19. Kerr WE. et al. Observações sobre a arquitetura dos ninhos e comportamento de algumas espécies de abelhas sem ferrão das vizinhanças de Manaus, Amazonas (Hymenoptera : Apoidea). Atas. Simp. Biota Amazônica, Manaus, AM, n. 5 (Zoologia), p. 255-309, 1967.
20. Laroca S, Almeida MC. Coexistência entre abelhas sem ferrão e formigas: Ninho de Paratrigona myrmecophila (Apidae) construído em ninho de Camponotus senex (Formicidae). Revta. Bras. Zool., Curitiba, PR.1989;6(4):671-680.
21. Imperatriz-Fonseca VL. et al. Subterranean nest structure of a stingless bee (Paratrigona subnuda Moure) (Meliponinae, Apidae, Hymenoptera). Ciência e Cultura, São Paulo, SP. 1972;24(7):662-666.
22. Camargo JMF. Biogeografia de Meliponini (Hymenoptera, Apidae, Apinae): a fauna Amazônica. In: ENCONTRO SOBRE ABELHAS, 1, 1994, Ribeirão Preto - SP, Anais... São Paulo, SP: USP, 1994, p. 46:59.
23. Nogueira-Neto P. et al. Biologia e manejo das abelhas sem ferrão. São Paulo, SP: Edição Tecnapis, 1986, 54p.
24. Vossler GG. Flower visits, nesting and nest defence behaviour of stingless bees (Apidae: Meliponini): suitability of bee species for meliponiculture in the Argentinean Chaco region. Apidologie. 2012;43:139-161.
25. Abraçado L. et al. "Magnetic material in head, thorax, and abdomen of Solenopsis substituta ants: A ferromagnetic resonance study", Journal of Magnetic Resonance. 2005;175(2):309-316.
26. Walker M. "A model for encoding of magnetic field intensity by magnetite-based magnetoreceptor cells", Journal of Theoretical Biology. 2008;250(1):85-91.
27. Begal S. et al. "Magnetic alignment in mammals and other animals", Mammalian Biology.2013;78(1):10-20.
28. Anderson JB, Vander Meer RK. Magnetic orientation in the fire ant, Solenopsis invicta. Naturwissenschaften 80, 568-570. 1993.
29. Menezes H. Própolis: uma revisão dos recentes estudos de suas propriedades farmacológicas. Arquivos do Instituto Biológico. 2005;72(3):405-411.
30. Bruening H.; Abelha Jandaíra. 3a ed. Natal: SEBRAE/ RN, 2006.
31. Serra BD. V. Abundância, distribuição espacial de ninhos de abelhas Meliponina (Hymenoptera, Apidae, Apini) e espécies vegetais utilizadas para nidificação em áreas de cerrado do Maranhão. Iheringia, Série Zoologia. 2009;99(1):12-17.
32. Kerr W. E. et al. Abelha Uruçu : Biologia, Manejo e Conservação – Belo Horizonte-MG: Acangaú, 1996. 144 p.: il., (Coleção Manejo da vida silvestre; 2).
33. Rosso L. J. M. et al. Meliponicultura em Brasil I: situacion em 2001 y perspectivas. In: Seminário Mexicano sobre Abejas sin Aguijón, 2., Yucatán, 2001. Anais... Yucatán, 2001. p. 28-35.
34. Campos LAO, Peruquetti RC. Biologia e criação de abelhas sem ferrão. Informe Técnico. Viçosa: UFV. 1999; n. 82, 38p.
35. Camargo JMF, Pedro SRM. Meliponini Lepeletier, 1836. In Moure, J. S., Urban, D., Melo, G. A. R. (Orgs). 2013. Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region - online version. Disponível em: http://www.moure.cria.org.br/catalogue. Acessado em 05/07/2017
36. Waldschmidt AM. Meliponicultura na Bahia. In: Congresso Baiano de Apicultura, 2. 2002, Paulo Afonso, BA. Anais... Paulo Afonso: p. 166-168. 2002.
37. Sakagami SF. Stingless bees. In: Herman, H. R. ed. Social Insects III. London, Academic Press, p. 361-423, 1982.
38. AQUINO IS. Abelhas Nativas da Paraíba. 1ª edição. João Pessoa. Editora Universitária - UFPB. 2006. 91p.
39. Lorenzon MCA. et al. Flora visitada pelas abelhas eussociais (Hymenoptera, Apidae) na Serra da Capivara, em caatinga do Sul do Piauí. Neotropical Entomology. 2003;32(1):27-36.
40. Martins CF.et al. Espécies arbóreas utilizadas para nidificação por abelhas sem ferrão na caatinga (Seridó, PB; João Câmara, RN). Biota Neotropica. 2004;4(2):1-8.
41. Antonini Y. Efeitos de variáveis ecológicas na ocorrência de Melipona quadrifasciata (Apidae, Meliponini) em fragmentos urbanos e rurais. 2002. Tese de doutorado, Universidade Federal de Minas Gerais, Belo Horizonte.
42. Ferreira R.LC, Vale AB. Subsídios básicos para o manejo florestal da caatinga. Anais do 2º Congresso Nacional sobre Essências Nativas:368-375. 1992.
43. Villas-Bôas J. Manual Tecnológico: Mel de Abelhas sem Ferrão. Brasília – DF. Instituto Sociedade, População e Natureza (ISPN). Brasil, 2012.
44. Werneck HA.; Faria-Mucci GM. Abelhas sem Ferrão (Hymenoptera:Apidae, Meliponini) da Estação Ecológica de Água Limpa, Cataguases-MG, Brasil. EntomoBrasilis. 2014;7(2):164-166.
45. Pedro SRM.; Camargo JM. F. Meliponini Neotropicais: o gênero Partamona Schwarz, 1939 (Hymenoptera, Apidae). Revista Brasileira de Entomologia, 47(supl. 1): 1-117. 2003.
46. Pioker-Hara FC. Determinantes da densidade e distribuição de ninhos e diversidade de espécies de meliponineos (Apidae, Meliponini) em áreas de cerrado de Itirapina, SP. Tese de doutorado em Ecologia. Universidade de São Paulo, USP, Brasil. 2011.
47. Lacerda DC. O. Influência da Orientação Magnética na Nidificação de Abelhas Nativas na Zona da Mata Pernambucana. 2017. Dissertação (Mestrado em Ciências Agrarias "Agroecologia"). Universidade Federal da Paraíba-UFPB, Bananeiras-PB, 77p.
48. Souza SGX. et. al. As abelhas sem ferrão (Apidae; Meliponina) residentes no campus Federação/Ondina da Universidade Federal da Bahia, Salvador. Candombá - Revista Virtual. 2005;1(1):57–69.
49. Silva FJA. Preferência de Nidificação em Abelhas Indígenas. 2016. Dissertação (Mestrado em Ciências Agrarias "Agroecologia"). Universidade Federal da Paraíba-UFPB, Bananeiras-PB, 90p.