
Resumo
Métodos clássicos de análise de modelos não lineares são amplamente utilizados em estudos da cinética de degradação ruminal. Como esse tipo de estudo envolve medidas repetidas na mesma unidade experimental, propõe-se o uso de modelos não lineares mistos (MNLM), buscando resolver os problemas de heterogeneidade de variâncias das respostas, de correlação entre as medidas repetida se a consequente falta de esfericidade da matriz de covariâncias. Os objetivos deste trabalho envolvem apresentar uma avaliação da aplicabilidade dos MNLM na estimação de parâmetros para descrever a cinética de degradação ruminal in situ da matéria seca de fenos de capim-Tifton 85 e comparar os seus resultados com os obtidos da análise usual realizada em duas fases. Os novilhos utilizados no ensaio foram alimentados com rações compostas por três diferentes combinações de volumoso e concentrado e dois fenos com diferentes qualidades nutricionais. A abordagem proposta mostrou-se tão efetiva quanto à tradicional para a estimação dos parâmetros do modelo. Contudo, ela modela de forma adequada a correlação entre os dados longitudinais, o que pode afetar as estimativas obtidas, o erro padrão associado a elas e, potencialmente, alterar os resultados das inferências. É bastante atraente quando a pesquisa busca entender o comportamento do processo da degradação dos alimentos ao longo dos tempos de incubação.
Palavras-chave: Cinética de degradação ruminal; Dados longitudinais; Efeitos aleatórios; Matéria seca; Matriz de variâncias e covariâncias.
Abstract
Classical methods of analysis of nonlinear models are widely used in studies of ruminal degradation kinetics. As this type of study involves repeated measurements in the same experimental unit, the use of mixed nonlinear models (MNLM) is proposed, in order to solve problems of heterogeneity of variances of the responses, correlation among repeated measurements and consequent lack of sphericity in the covariance matrix. The aims of this work are to present an evaluation of the applicability of MNLM in the estimation of parameters to describe the in situ ruminal degradation kinetics of the dry matter of Tifton 85 hay and to compare the results with those obtained from the usual analysis in two-phases. The steers used in the trial were fed diets composed of three different combinations of roughage and concentrate and two hays with different nutritional qualities. The proposed approach was proven as effective as the traditional one for estimating model parameters. However, it adequately models the correlation among the longitudinal data, which can affect the estimates obtained, the standard error associated with them and potentially change the results of the inferences. It is quite attractive when the research seeks to understand the behavior of the process of food degradation throughout the incubation times.
Keywords: Ruminal degradation kinetics; Longitudinal data; Covariance matrix; Random effects; Dry matter.
Seção: Zootecnia
Recebido
9 de março de 2019.
Aceito
7 de maio de 2019.
Publicado
16 de junho de 2020.
www.revistas.ufg.br/vet
Como citar - disponível no site, na página do artigo.
Introdução
O conhecimento do processo de degradação dos alimentos pelos microrganismos do rúmen é importante em estudos de avaliação de alimentos para ruminantes, pois o conhecimento do valor nutritivo potencial dos alimentos, por meio da degradação ruminal, permite o seu emprego racional como alimento único ou como ingrediente de misturas mais complexas(1,2).
O consumo alimentar é altamente correlacionado com a sua composição nutricional e com a digestibilidade, uma vez que a regulação fisiológica ocorre quando há o aumento de consumo de matéria seca com o aumento da digestibilidade, corroborando a saciedade do animal(3,4).
Dentre as técnicas empregadas para avaliar a degradação ruminal dos alimentos, a técnica in situ tem sido a mais extensivamente utilizada, a qual consiste em determinar o desaparecimento de componentes da amostra de alimentos acondicionados em sacos de náilon incubados no rúmen, por períodos variáveis(5). Embora não permita que o alimento sofra todos os eventos digestivos, como mastigação e ruminação, de acordo com Pereira et al.(6), o extenso uso dessa técnica está ligado à sua rápida e fácil execução, pois requer uma pequena quantidade de amostra do alimento-teste, possibilita sua exposição ao ambiente ruminal, além dos seus resultados serem próximos aos encontrados com ensaio in vivo.
Os modelos não lineares são amplamente utilizados em estudos que buscam estimar os parâmetros da cinética de degradação ruminal in situ(7). Porém, devido à grande quantidade de fatores envolvidos na realização desses experimentos, diferentes procedimentos e modelos de análises podem ser utilizados(6,8-12). Como esses estudos envolvem dados longitudinais, pois as medidas de degradação são obtidas sistematicamente ao longo do tempo nas mesmas unidades experimentais, espera-se que exista correlação não nula entre as medidas sucessivas e heterogeneidade das variâncias entre as medidas feitas nas diversas ocasiões. Esses aspectos não são levados em conta nos métodos clássicos de estimação e de análise univariada de variância, o que pode alterar os resultados das inferências feitas sobre os parâmetros do modelo. Pasternak e Shalev(13) afirmaram que o simples ajuste de uma regressão não linear a dados longitudinais pode ser ineficiente, uma vez que não considera a heterogeneidade de variâncias. Sartorio(14) e Carvalho et al.(15), dentre outros, afirmaram que a análise tradicional de dados realizada em duas fases(5,8,9) também deve ser ineficiente, porque, além de não considerar a heterogeneidade das variâncias, também não leva em conta a possível correlação entre as medidas repetidas ao longo do tempo, o que fere a premissa da esfericidade da estrutura de variâncias e covariâncias, que é exigência dos modelos de regressão clássicos. Uma abordagem mais atual para análise de dados longitudinais com comportamento não linear para as respostas médias envolve o uso de modelos não lineares mistos (MNLM), que ainda são pouco empregados na análise de ensaios de degradabilidade ruminal(14). Diante do exposto, o objetivo deste trabalho é apresentar e avaliar a aplicabilidade de modelos não lineares mistos para descrever a cinética de degradação ruminal in situ de fenos de capim-Tifton 85, em novilhos alimentados com rações compostas por diferentes combinações de volumoso:concentrado e fenos de duas diferentes qualidades nutricionais, comparando os seus resultados com os obtidos da análise tradicional, que é realizada em duas fases.
Materiais e métodos
Os dados foram obtidos do experimento desenvolvido por Feitosa et al.(17), na Unidade Animal de Estudos Digestivos e Metabólicos do Departamento de Zootecnia da Faculdade de Ciências Agrárias e Veterinárias/UNESP, Campus de Jaboticabal, no período de maio de 2001 a dezembro de 2002.
Foram utilizados seis novilhos mestiços (Bos taurus × Bos taurus indicus) canulados no rúmen, com peso médio de 550±35kg e aproximadamente três anos e meio de idade. Durante o período experimental, os animais foram mantidos em baias individuais, providos de bebedouro e comedouro, recebendo dietas compostas de feno de capim-Tifton 85 (TIF) e concentrado em duas refeições diárias, às 7h30 e às 18h30. A quantidade de alimento fornecida foi estabelecida em função do consumo máximo diário verificado com a dieta de maior porcentagem do volumoso e de pior qualidade nutricional. Os animais foram adaptados aos tratamentos durante 14 dias, seguidos de 10 dias de coleta de amostras para cada período experimental.
As seis rações formuladas foram compostas pela combinação de três diferentes relações,70:30, 50:50 e 30:70 de volumoso (V) e concentrado(C), com base na matéria seca, e dois fenos de capim-Tifton 85 com 4% (TIF4) e 10% (TIF10) de proteína bruta, ambos picados em partículas de 5 mm. O concentrado foi composto de casca de soja (Glycine max L.), milho (Zea mays L.) moído e farelo de girassol (Helianthus annuus L.). Os tratamentos foram identificados como: V70TIF4, V70TIF10, V50TIF4, V50TIF10, V30TIF4 e V30TIF10.
A degradabilidade ruminal da matéria seca (MS) foi determinada pela técnica in situ, usando-se sacos confeccionados em náilon, medindo aproximadamente 14×7 cm, com poros de 50µm, contendo amostras de cada uma das dietas experimentais moídas com granulometria de 5 mm. Foram incubados 4,5g de amostra (base na MS), amarrados em elos de uma corrente de aço para todos os tempos de incubação. Os dez tempos de incubação no rúmen foram de 120, 96, 84, 72, 60, 48, 24, 12, 6 e 3 horas com duas réplicas para cada horário e animal, adotando-se o sistema de colocação dos sacos de náilon nos tempos de incubação e retirada simultânea de todos os sacos ao final do período.
Após a incubação, os sacos de náilon foram pré-lavados em água corrente para a retirada do excesso de conteúdo ruminal e colocados em água com gelo por 20 min. Em seguida, foram lavados durante 15 minutos em uma máquina do tipo tanquinho (sendo que a água da máquina foi trocada a cada cinco minutos). Após esta etapa, os sacos com os resíduos foram secos em estufa a 65°C, com circulação forçada de ar durante 48 horas.
Os sacos com os resíduos secos foram pesados e seus resíduos moídos em moinho de facas, com peneira de 1mm de diâmetro, armazenados em frascos com tampa e identificados. As amostras foram secas em estufa regulada a 105°C por 12horas e as análises para determinação da matéria seca foram realizadas no Laboratório de Ingredientes e Gases Poluentes(LIGAP) do Departamento de Zootecnia da FCAV/UNESP, Campus de Jaboticabal – SP.
O experimento básico foi planejado em quadrado latino 6×6 com seis animais em seis períodos experimentais distintos, recebendo um dos seis tratamentos (V70TIF4, V70TIF10, V50TIF4, V50TIF10, V30TIF4 e V30TIF10). Em todas as unidades experimentais, foram feitas as medidas repetidas ao longo do tempo. Dentre as características avaliadas nas amostras, a variável resposta considerada nesse trabalho foi a porcentagem de desaparecimento da matéria seca (%MS).
Admitiu-se que o comportamento não linear da porcentagem de desaparecimento da matéria seca (%MS) ao longo dos tempos de incubação pode ser bem explicado pelo modelo de Orskov e McDonald(16). Assim, a %MS avaliada no instante xij, no i-ésimo animal que recebeu o κ-ésimo tratamento no j-ésimo período, para i, j, κ = 1, 2, …, 6, pode ser expressa por:
yijκ = β1κ + β2κ[1 - exp(-β3κxij)] + εijκ (1)
em que β1κ representa a fração rapidamente solúvel das parcelas que receberam o tratamento κ, β2κ é a fração que pode ser degradada, se houver tempo, das parcelas que receberam o tratamento κ, β3κ é a taxa de degradação da fração das parcelas que receberam o tratamento κ e εijκ é o erro experimental associado à observação yijκ.Admitiu-se que εijκ ~ N(0, σ²ε ), ou seja, as medidas repetidas nas mesmas unidades experimentais são independentes e a variância é constante em todos os tratamentos e períodos.
Do modelo (1) derivam parâmetros importantes para o balanceamento de rações, como a degradabilidade potencial, definida como PD = β1 + β2, e a degradabilidade efetiva (ED) que é calculada como: DE5% = β1+(β2β3)/(β3+c), em que c = 5%/hora é a taxa de passagem das partículas no rúmen para animais com médio consumo.
A metodologia denominada análise usual em duas fases foi a mesma utilizada nos trabalhos de Santoro et al.(8); Teixeira et al.(5) e Jobim et al.(9), dentre outros. Na primeira fase, ajustou-se o modelo de Orskov e McDonald(16) para os dados de %MS de cada unidade experimental e com as estimativas dos seus parâmetros (β1κ, β2κ, β3κ) criou-se uma nova planilha de dados. Na segunda fase, essas estimativas foram utilizadas como variáveis respostas em uma série de três análises de variância univariadas (ANOVA) independentes tendo como covariáveis os fatores associados ao delineamento experimental (Animal, Período e o fator de Tratamento), que até o momento não foram incorporados na análise. Desta forma tem-se, para , u = 1, 2, 3 o modelo:
β = μ + Aui + Puj + Tuκ + εijκ (2)
em que μ é uma constante comum a todas as observações; Aui é o efeito do i-ésimo animal na u-ésima variável resposta; Puj é o efeito do j-ésimo período na u-ésima variável resposta; Tuκ é o efeito do κ-ésimo tratamento na u-ésima variável resposta e εijκ é o erro experimental associado à u-ésima variável medida no i-ésimo animal, que no j-ésimo período que recebeu o tratamento κ, para i, j, κ = 1, 2, …, g e g é o número de tratamentos envolvidos no ensaio. Quando a hipótese de igualdade das médias dos tratamentos for rejeitada (p < 0,05), elas podem ser comparadas utilizando um teste de comparações múltiplas, como o teste de Tukey, por exemplo.
A análise utilizando o modelo não linear misto (MNLM) foi realizada como descrita por Lindstrom e Bates(18) e utilizada por Gómez, Muñoz e Betancur(19).De acordo com Lindstrom e Bates(18), o MNLM pode ser escrito como:
yi = f(Xi, β, Zi, bi ) + εi (3)
em que yi = [yi1, yi2,...,yini ] é o vetor (ni×1) de respostas do i-ésimo indivíduo, sendo ni o número de tempos observados no i-ésimo indivíduo; f(Xi, β, Zi, bi) é um vetor (ni×1) de funções de resposta não linear; Xi = [xi1,..., xiw ] é uma matriz (ni×W) de valores intraindivíduos, podendo incluir somente os instantes de avaliação; β é um vetor (p×1) de parâmetros populacionais desconhecidos; Z é uma matriz (ni×q) de posto completo, de constantes conhecidas que associa yi a bi, que é um vetor (q×1) de coeficientes aleatórios de regressão, não observados e é um vetor (q×1) de erros aleatórios intraindivíduo. É comum assumir que as observações feitas em diferentes indivíduos são independentes entre si e que εi ~ Nni (0,Ri), sendo Ri a matriz de variâncias e covariâncias de dimensão (ni×ni), que depende de somente pela sua dimensão. Apesar de se admitirem diversas aplicações que Ri = σ²Ini, onde Ini é uma matriz identidade de dimensão ni , Ri, pode assumir qualquer estrutura de covariâncias marginal especial, como a autorregressiva de primeira ordem (AR(1)), por exemplo. Admite-se que os efeitos aleatórios, bi, são independentes e identicamente distribuídos, bi ~ Nq(0,σ²D), em que σ²D é uma matriz de variâncias e covariâncias, cuja dimensão depende do número de efeitos aleatórios considerados no modelo. Admite-se ainda que os bi são independentes dos εi.
Para explicar o comportamento da digestibilidade média da MS (%), por tratamento, utilizou-se a função não linear apresentada em (1), para i, j, κ = 1, …, 6. Os animais fistulados avaliados em períodos distintos foram considerados unidades experimentais diferentes. Como os dados em análise são completos e balanceados em relação ao tempo, tem-se inicialmente, porque ainda não se sabe quantos e onde serão incluídos os efeitos aleatórios, que: ni = 10 tempos, xi = 3, 6, 12, 24, 48, 60, 72, 84, 96 e 120 e Ri = R = σ²I , é comum a todos os perfis individuais de resposta.
Para sugerir em quais parâmetros é conveniente incluir um efeito aleatório, foram confeccionados os gráficos de perfis individuais de respostas e de intervalos de 95% de confiança para cada um dos parâmetros do modelo não linear, utilizando as estimativas dos parâmetros dos ajustes feitos para todos os indivíduos. Uma variabilidade incomum dos pontos em alguma parte do gráfico de perfis individuais de resposta (nos pontos iniciais ou finais do processo, ou na curvatura dos perfis, por exemplo), associada a algum(ns) parâmetro(s) do modelo não linear escolhido, sugere a inclusão de efeito aleatório nesse(s) parâmetro(s). Já no gráfico de intervalos de 95% de confiança, a não sobreposição dos intervalos calculados para certo parâmetro indica a inclusão de um efeito aleatório a este parâmetro(20).
O modelo (1) com efeito aleatório associado à fração rapidamente solúvel, β1, pode ser escrito como:
yik = (β1k + b1i ) + β2k[1- exp(-β3kxi)] + εijκ (4)
em que b1i é o efeito aleatório associado a β1. Neste exemplo, diz-se que o efeito aleatório está associado de forma linear ao parâmetro de efeito fixo β1, como descrito por Hist et al.(21) e Vonesh e Carter(22). Se o efeito aleatório está associado ao parâmetro β2, diz-se que o mesmo ocorrerá de forma linear. Porém, quando um efeito aleatório está associado ao parâmetro β3, este ocorrerá na forma não linear, como proposto por Lindstrom e Bates(18).
Se essas técnicas informais de escolha de efeitos aleatórios não forem conclusivas, podem ser comparados todos os possíveis modelos, utilizando testes de razão de verossimilhanças (TRV) em modelos encaixados ou os critérios de informação de Akaike (AIC) ou bayesiano (BIC), quando os modelos não forem encaixados. Neste caso, o modelo que apresentou o menor valor dessas estatísticas (AIC ou BIC) foi considerado o mais adequado(20).
Quando o modelo envolveu mais de um efeito aleatório, escolheu-se uma estrutura para a matriz D, associada ao vetor de efeitos aleatórios, que seja parcimoniosa e consiga explicar bem a variabilidade e as covariâncias entre tais efeitos.
Para o modelo final e após a escolha dos efeitos aleatórios, foram ajustadas curvas médias distintas para cada tratamento e as comparações entre tratamentos foram feitas como descrito em Sartorio(14). Todas as análises foram realizadas utilizando o pacote nlme do software R(23), considerando um nível de significância α= 0,05 em todos os testes de hipóteses.
Resultados e Discussão
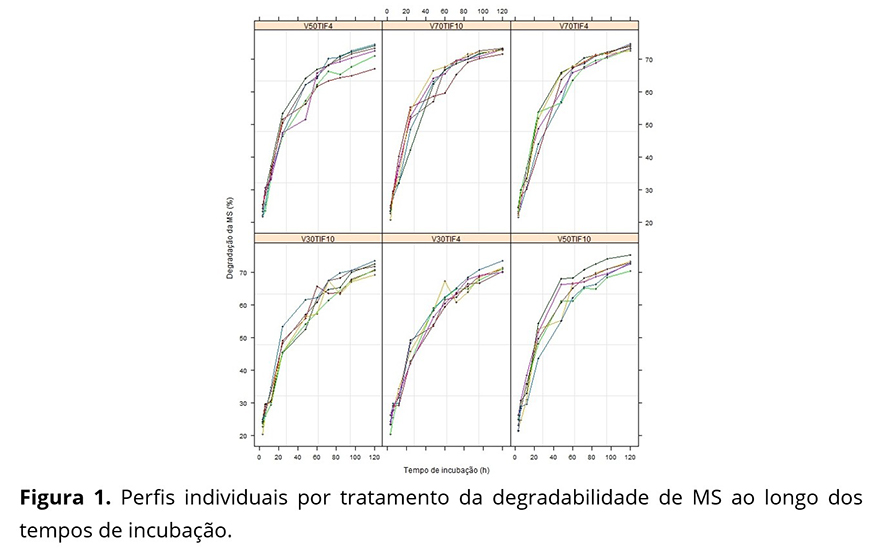
Os perfis individuais da degradabilidade da matéria seca (MS) ao longo do tempo, por tratamento (Figura 1) têm comportamentos semelhantes, apresentam certa heterogeneidade de variâncias ao longo do tempo e indicam que o modelo não linear (1) pode explicar bem a relação entre a porcentagem de degradação da MS e os tempos de incubação.
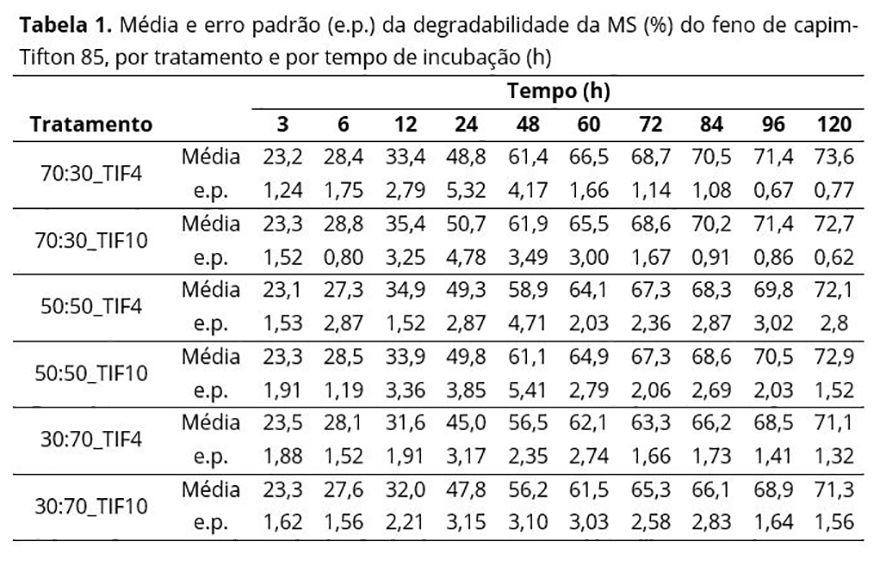
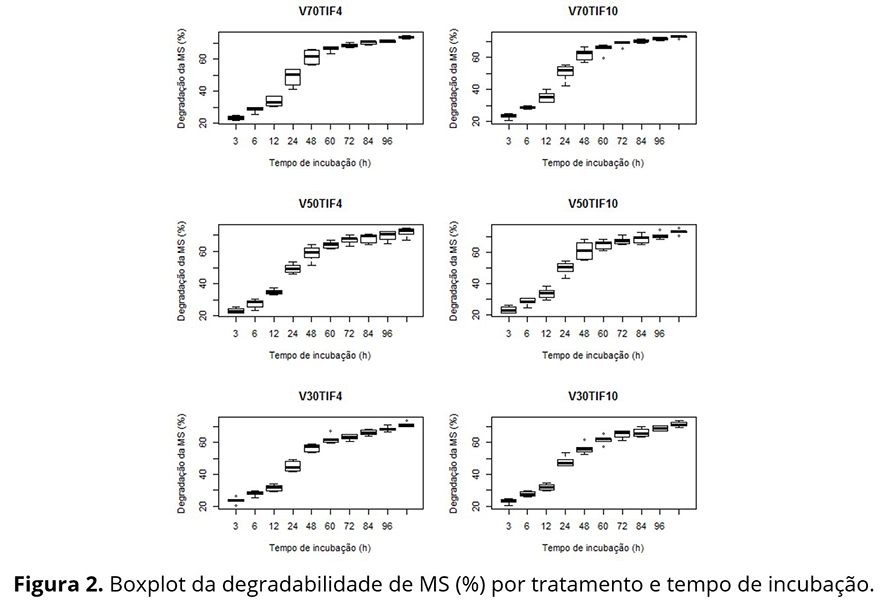
Na Tabela 1 nota-se, de um modo geral, um aumento da variabilidade das respostas até as 24 ou 48 horas de incubação e uma posterior diminuição até 120 horas, para todos os tratamentos, aspecto não tão evidente nos gráficos de perfis individuais (Figura 1), sendo mais evidente na Figura 2.
O modelo de Orskov e McDonald(16) foi ajustado para cada um dos trinta e seis indivíduos, resultantes da combinação dos seis animais avaliados em seis diferentes períodos. Para as três variáveis respostas estudadas (estimativas de β1, β2, e β3), os testes de Shapiro-Wilk e de Bartlett confirmaram a normalidade dos erros (p > 0,05) e a homogeneidade de variâncias entre os seis tratamentos ( p > 0,05), respectivamente.
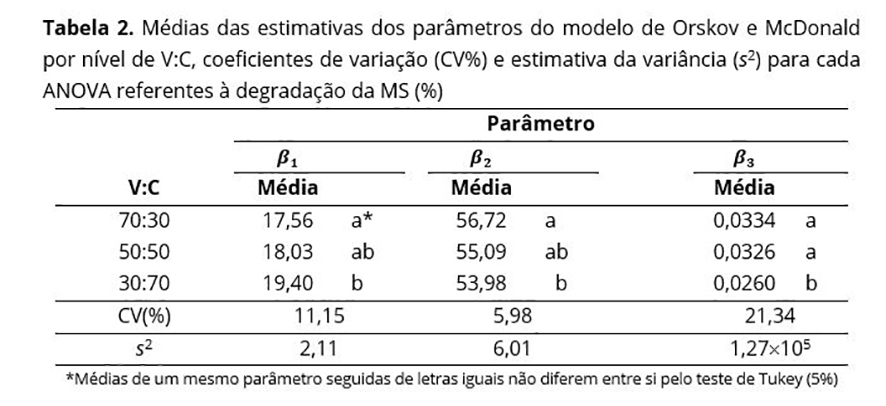
As ANOVAs individuais dessas variáveis respostas indicaram que não houve interação ( p > 0,05) entre os níveis dos fatores V:C e TIF, que não se observou efeito do fator TIF na degradabilidade da MS, ocorrendo diferenças significativas (p < 0,05) somente entre as médias dos níveis do fator V:C. Esses resultados concordaram com aqueles obtidos por Feitosa et al.(17).
Os coeficientes de variação (Tabela 2) foram considerados médio-alto para os parâmetros β1 e β3, e baixo-médio para o parâmetro β2, segundo a classificação de Vaz et al.(24). Pelo teste de Tukey, percebeu-se que os animais que receberam 70% de volumoso na ração apresentaram maior valor de β2 e menor valor de β1 que os animais que receberam somente 30% de volumoso. Os tratamentos com 70 e 50% de volumoso apresentaram valores médios de β3 iguais entre si e superiores ao do tratamento com 30% de volumoso.
Os intervalos de 95% de confiança construídos para os três parâmetros a partir dos ajustes do modelo para os 36 perfis individuais de degradabilidade da MS (%) (Figura 3), sugerem a inclusão de efeito aleatório em β2, β3 e, possivelmente, em β1, pois o número de intervalos não sobrepostos para este parâmetro é pequeno. Para confirmar essas sugestões, sete modelos com efeitos aleatórios foram sugeridos (Tabela 3).
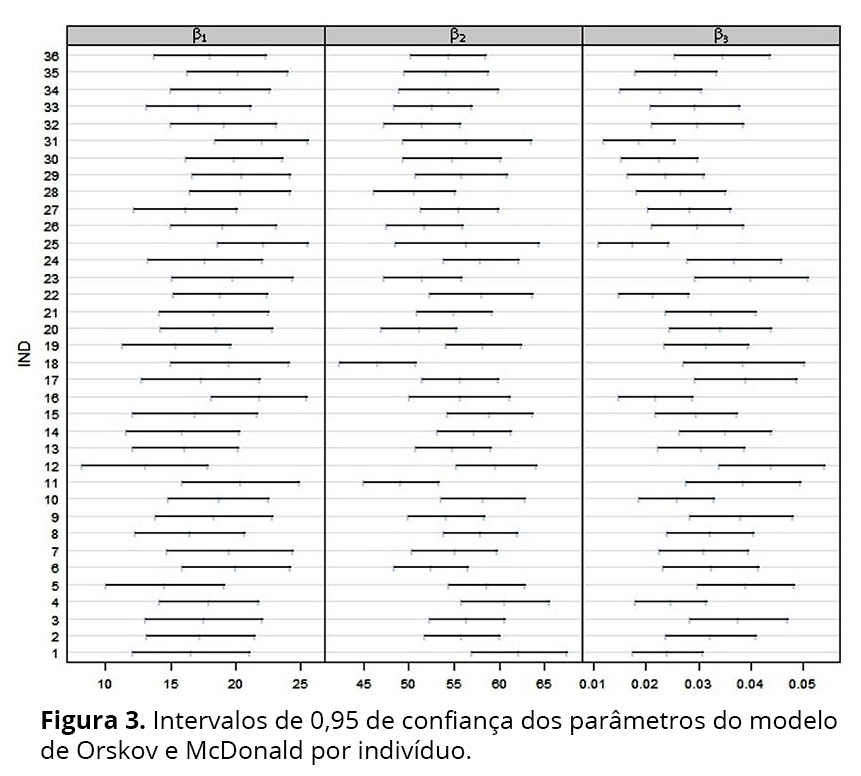

Para buscar a estrutura de variâncias e covariâncias adequada para os efeitos aleatórios, combinaram-se os sete modelos com efeitos aleatórios apresentados na Tabela 3, com quatro estruturas para a matriz D(pdSymm: matriz geral positiva definida, sem estrutura adicional; pdLogChol: matriz geral positiva definida, sem estrutura adicional, usando uma parametrização log-Cholesky; pdDiag: diagonal; pdIdent: múltiplo de uma identidade), admitindo Ri = σ²Ini.
Testes da Razão de Verossimilhanças (TRV) e os critérios AIC e BIC indicaram que o modelo MR6, com efeitos aleatórios em β2 e β3, com matriz não estruturada para D, proporcionou melhor ajuste que o modelo com efeitos fixos (MF0) e todos os demais modelos não lineares mistos.
Também foi testada a necessidade de utilizar outra estrutura para a matriz R (autorregressiva de primeira ordem – AR(1); autorregressiva de primeira ordem com heterogeneidade de variâncias – ARH(1) ou simetria composta – SC), mas os resultados confirmaram que a estrutura R = σ²I10 é a mais adequada.
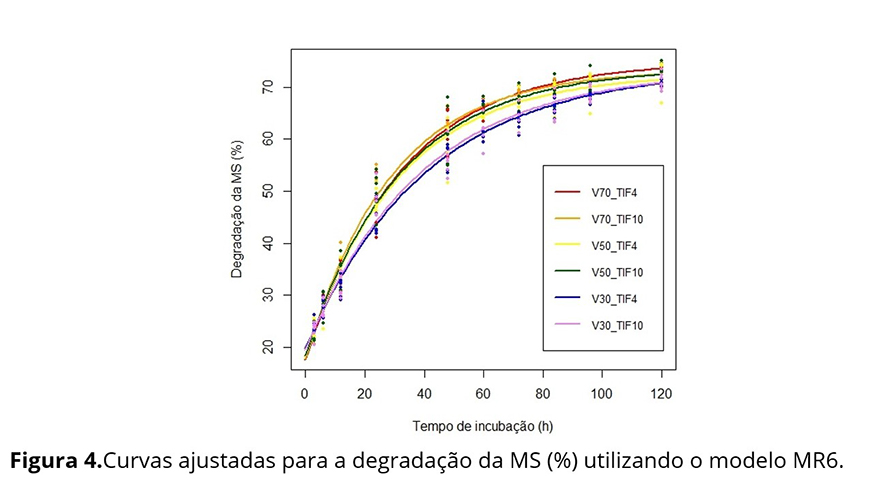
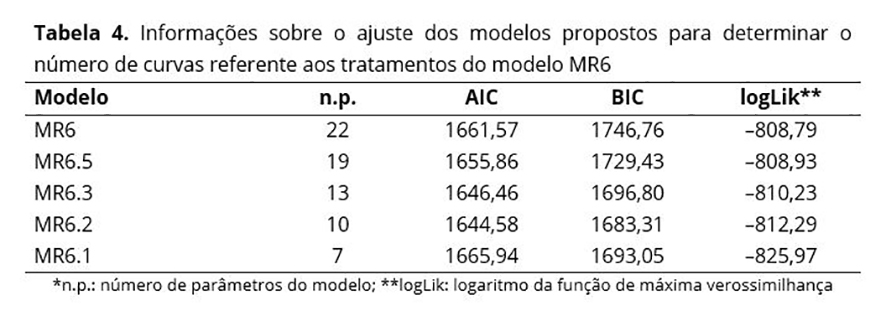
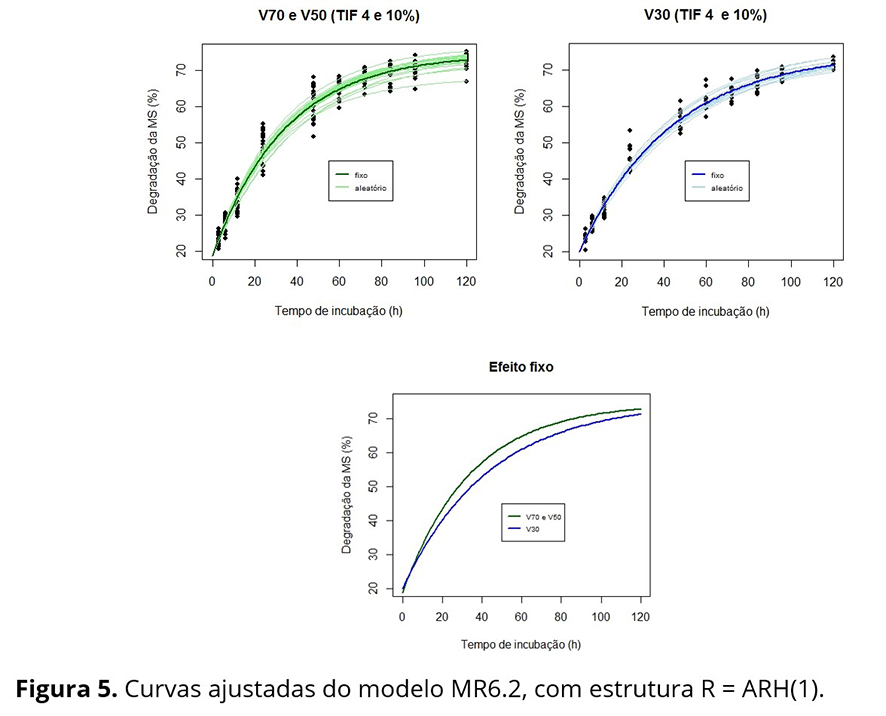
Na Figura 4, que apresenta as curvas médias ajustadas aos seis tratamentos, observa-se que as curvas dos tratamentos V30TIF4 e V30TIF10 são muito semelhantes, assim como as curvas dos demais tratamentos. Considerando a estrutura aleatória do modelo MR6 e as curvas médias apresentadas, foram propostos alguns modelos alternativos: i) MR6.5: cinco curvas distintas, sendo uma única curva para os tratamentos V30TIF4 e V30TIF10 e uma curva para cada um dos outros tratamentos; ii) MR6.3: três curvas distintas, sendo uma para os tratamentos V70TIF4 e V70TIF10, outra para V50TIF4 e V50TIF10 e outra para V30TIF4 e V30TIF10; iii) MR6.2: duas curvas distintas, sendo uma para os tratamentos V30TIF4 e V30TIF10 e outra para os demais tratamentos V70TIF4, V70TIF10, V50TIF4 e V50TIF10; iv) MR6.1: uma única curva para todos os perfis médios. Os modelos foram comparados utilizando TRV e os critérios AIC e BIC e concluiu-se que o modelo MR6.2 com 10 parâmetros é tão bom quanto os demais modelos, que envolvem 13, 19 ou 22 parâmetros (Tabela 4). Logo, as curvas de degradabilidade da MS (%) em função dos tempos de incubação foram consideradas idênticas para as porcentagens de volumoso de 70 e 50, sendo apenas a porcentagem de 30 considerada diferente das demais. A qualidade nutricional dos fenos de capim-Tifton 85 (TIF4 e TIF10) não influenciou na degradabilidade da MS (%).
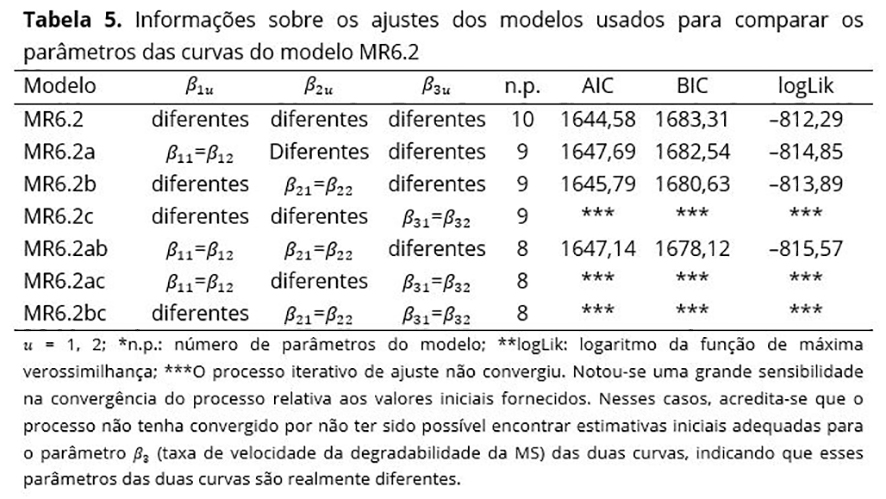
Os parâmetros do modelo MR6.2 também foram comparados utilizando os modelos apresentados na Tabela 5. Testes da razão de verossimilhanças foram realizados e permitiram concluir que o modelo MR6.2 com duas curvas e parâmetros distintos foi o mais indicado para a descrição da variável resposta em estudo.
Lembrando que a estrutura aleatória do modelo foi selecionada anteriormente à dos efeitos fixos, Pinheiro e Bates(20) recomendam conferir a estrutura da matriz R, ou seja, se essa estrutura permanece a mesma escolhida antes de definir os efeitos fixos. Dentre as estruturas mais complexas (AR(1), ARH(1), e CS) comparadas, por meio de TRV, com R = σ²I10 , a estrutura ARH(1), com variâncias diferentes nos diversos tempos de incubação e autocorrelação de primeira ordem, foi considerada mais adequada.
Os tratamentos com 70 ou 50% de volumoso não apresentaram diferenças entre si para todos os parâmetros do modelo, mas foram diferentes dos tratamentos em que a porcentagem de volumoso foi de 30% (também para todos os parâmetros do modelo). O modelo não linear misto final explica praticamente toda a variabilidade presente nos dados (Figuras 5 e 6), o que não ocorre com o modelo de duas fases.
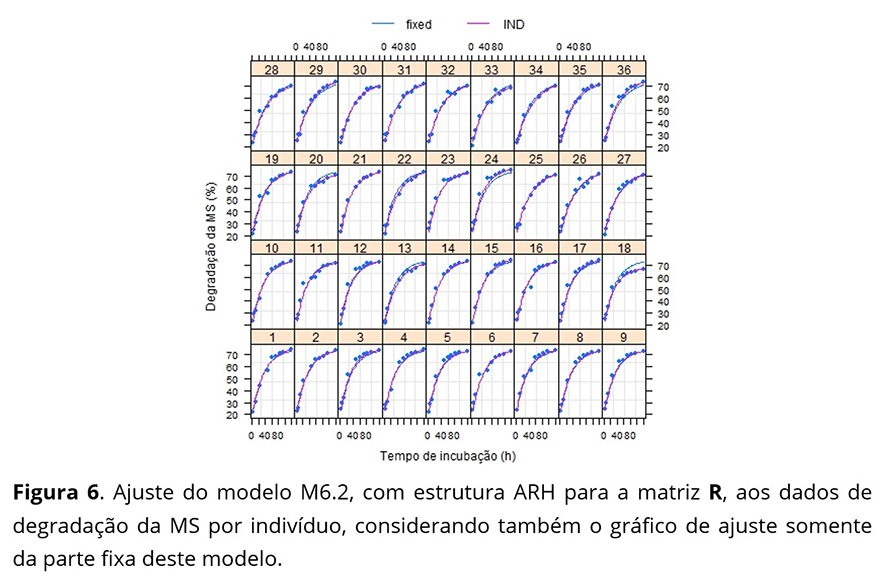
Na Figura 6 percebe-se um comportamento das respostas do indivíduo 18 bem distinto dos outros indivíduos, apresentando a maior variação referente à fração potencialmente degradável (β2). Este fato explica a necessidade de um maior valor predito do efeito aleatório, referente ao parâmetro β2, encontrado para o indivíduo 18.
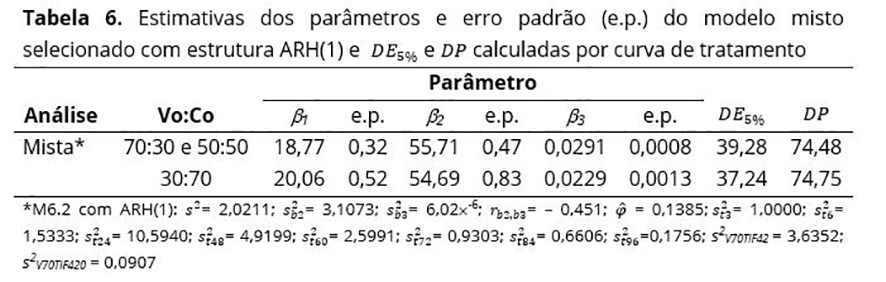
A menor proporção de concentrado proporcionou uma DE5% maior do feno de capim-Tifton, em relação às outras proporções consideradas. Já as DPs de ambos os tratamentos apresentaram praticamente os mesmos valores (Tabela 6), que foram condizentes aos obtidos por Jobim et al.(9).
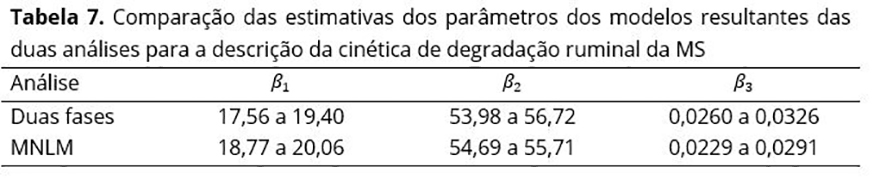
Nas duas abordagens, as porcentagens 70 e 50 de volumoso na ração não apresentaram diferença para nenhum dos parâmetros avaliados, ou seja, para a fração rapidamente degradável (β1), para a fração potencialmente degradável (β2) e para a taxa de degradação da fração potencialmente degradável (β3). Os valores das estimativas dos parâmetros β1 , β2 e β3 dos modelos finais da abordagem mista estão próximos das faixas de variação (ou dos intervalos de variação) obtidos com a abordagem de duas fases (Tabelas 6 e 7), tendo menor variação a abordagem mista.
Considerando o modelo final, MR6.2 com estrutura ARH(1) para a matriz R, a menor proporção de volumoso na ração proporcionou a menor DE5% do feno de capim-Tifton 85 (37,24%), em relação às outras proporções consideradas, estando próxima dos valores encontrados por Assis et al.(26), Balieiro e Melloti(25), como também próxima ao intervalo de valores obtidos na abordagem de duas fases (37,68 a 40,01) e mista (37,31 a 40,38). Já as DPs de ambos os tratamentos apresentaram valores muito próximos, estando também de acordo com os obtidos pela de duas fases (73,12 a 74,28), valores próximos aos obtidos por Jobim et al.(9) e superiores aos encontrados por Balieiro e Melloti(25). Além disso, a estimativa da variabilidade residual (s²) referente à análise de duas fases sofreu uma grande redução (de 6,84 para 2,02) já esperada, pois a variabilidade residual, que antes era explicada por uma única fonte de variação no modelo não linear clássico (análise de duas fases), passou a ser composta por variações entre indivíduos (variação residual mais efeitos aleatórios) e intraindivíduos (relativa à heterogeneidade de variâncias nos diversos tempos de incubação) no modelo não linear misto, justificando a sua redução. Dessa forma, o uso do MNLM se apresenta melhor que o método em duas etapas para diferenciar a degradabilidade efetiva e potencial de alimentos, pois, se o erro padrão das estimativas dos parâmetros é maior, os respectivos intervalos de 95% de confiança também serão maiores, diminuindo a probabilidade de encontrar diferença entre os tratamentos, ou seja, aumenta-se a probabilidade de aceitar erroneamente a hipótese de efeito nulo dos tratamentos (erro tipo II).
Os resultados referentes aos efeitos fixos obtidos com o uso de MNLM não foram distintos dos obtidos na análise de duas fases, nas condições deste ensaio. Contudo, a abordagem mista apresenta vantagens em relação à abordagem de duas fases, quando o interesse está na descrição do comportamento dos perfis individuais de respostas e na separação da variabilidade residual em fontes de variação entre e intraindivíduos.
Resultados semelhantes foram obtidos por Zanton e Heinrichs(27) que fizeram um estudo para a variável resposta fibra de detergente neutro (FDN), em que avaliaram três metodologias de análises de perfis de degradação alimentar in situ. Para isso, simularam 500 experimentos, considerando apenas quatro animais, dois períodos e oito tempos de incubação (72, 48, 24, 16, 8, 4, 2, e 1 hora). Nessas condições, os autores puderam concluir que, em muitos casos, o MNLM é tão bom ou melhor que a análise de duas fases.
A variabilidade provocada pelo delineamento experimental nas respostas foi bem explicada pelas estruturas de covariâncias utilizadas, indicando que a inclusão de parâmetros relacionados aos fatores de delineamento não seja necessária com o uso do modelo não linear misto.
Conclusão
O uso do modelo de Orskov e McDonald(16) com efeitos aleatórios proporcionou o melhor ajuste aos dados de degradabilidade de matéria seca, pois a variabilidade dos dados é mais bem descrita. O pacote nlme do software R foi bastante versátil no ajuste, possibilitando o ajuste e comparação de diversos modelos para a parte fixa, alternando diversas estruturas de variâncias e covariâncias.
O uso de modelos não lineares mistos na análise de dados de digestibilidade in situ é bastante vantajoso quando a pesquisa tem por objetivo entender o comportamento do processo de digestibilidade ao longo dos tempos de incubação. Se o interesse for restrito à estimação dos parâmetros do modelo não linear de Orskov e McDonald e ao cálculo das degradabilidades efetiva e potencial de dietas, pode-se usar qualquer uma das duas abordagens de análise apresentadas.
Apesar de esse estudo sugerir que, por reduzir a estimativa da variabilidade residual (s²) o uso do MNLM se apresenta melhor que o método em duas etapas para diferenciar a degradabilidade efetiva e potencial de alimentos, é necessário realizar simulações para comprovar tal afirmação.
Agradecimentos
Ao prof. Dr. José Valmir Feitosa por ceder os dados. À CAPES pela bolsa de estudos. Este trabalho é parte da tese de doutorado do primeiro autor, obtida pela ESALQ/USP.
Referências