DOI: 10.1590/1089-6891v20e-57228
MEDICINA VETERINÁRIA
ATIVIDADE ANTIMICROBIANA DO EXTRATO ETANOLICO DA Commiphora leptophloeos (MART.) J. B. GILLETT FRENTE A Staphylococcus spp. ISOLADOS DE CASOS DE MASTITE EM RUMINANTES
ANTIMICROBIAL ACTIVITY OF ETHANOLIC EXTRACTS FROM Commiphora leptophloeos (MART.) J. B. GILLETT AGAINST Staphylococcus spp. ISOLATED FROM CASES OF MASTITIS IN RUMINANTS
Isamara Ferreira da Silva¹ ORCID - http://orcid.org/0000-0001-7739-3664
Amanda Leite Guimarães¹ ORCID - http://orcid.org/0000-0001-8235-6024
Vinícius Santana Amorim¹ ORCID - http://orcid.org/0000-0001-8657-3202
Telma Maria Guedes da Silva² ORCID - http://orcid.org/0000-0001-5778-0384
Rodolfo de Moraes Peixoto3* ORCID - http://orcid.org/0000-0002-5757-5935
Xirley Pereira Nunes¹ ORCID - http://orcid.org/0000-0002-9714-8575
Tânia Maria Sarmento Silva² ORCID - http://orcid.org/0000-0003-1887-2598
Mateus Matiuzzi da Costa¹ ORCID - http://orcid.org/0000-0002-98842112
¹Universidade do Vale do São Francisco, Petrolina, PE, Brasil.
²Universidade Federal Rural de Pernambuco, Recife, PE, Brasil.
³Instituto Federal do Sertão Pernambucano, Petrolina, PE, Brasil.
*Autor para correspondência - rodolfo.peixoto@ifsertao-pe.edu.br
Resumo
Considerando o potencial terapêutico de plantas medicinais como alternativas à antibioticoterapia, a pesquisa visa avaliar a atividade antimicrobiana de Commiphora leptophloeos contra isolados de Staphylococcus spp. de leite de ruminante com mastite subclinica. Para isso, foi elaborado o extrato etanólico bruto da casca e folhas de Commiphora leptophloeos, sendo as mesmas caracterizadas quimicamente por HPLC-DAD-MS e por MALDI-TOF. Os extratos foram então avaliados quanto aos seus efeitos antimicrobianos contra 60 isolados de Staphylococcus spp pela técnica de microdiluição em caldo para determinação da concentração bactericida mínima. Além disso, os extratos foram avaliados quanto à sua capacidade de interferir na formação do biofilme e com o biofilme já consolidado. Apesar de todos os extratos testados terem apresentado ação antimicrobiana, valores menores de CBM foram registrados para o extrato da casca na concentração 781,2 μg/mL (25/60). Os extratos da casca e das folhas foram capazes de interferir nos estágios iniciais da formação do biofilme, mas não ocorreu interferência do extrato no biofilme consolidado. Houve alta sensibilidade do isolados testados quando submetidos a extratos de folhas e casca de Commiphora leptophloeos, bem como a capacidade de extratos interferirem na formação de biofilme, indicando seu potencial uso na terapia de mastite de ruminantes.
Palavras-chave: Mastite, Staphylococcus spp, Imburana de Cambão, Fitoterapicos, biofilme.
Abstract
Considering the therapeutic potential of medicinal plants as alternatives to antibiotic therapy, the research aims to evaluate the antimicrobial activity of Commiphora leptophloeos against isolates of Staphylococcus spp. from the milk of ruminants with subclinical mastitis.For this, the crude ethanolic extract from the bark and leaves of Commiphora leptophloeos was prepared, with these being chemically characterized by HPLC-DAD-MS and by MALDI-TOF. The extracts were then evaluated as to their antimicrobial effects against 60 isolates of Staphylococcus spp. through the broth microdilution technique to determine the minimum bactericidal concentration. In addition, the extracts were evaluated as to their ability to interfere with biofilm formation and with the already established biofilm. Although all tested extracts showed antimicrobial action, lower MBC values were recorded for the bark extract in the concentration 781.2 μg/mL (25/60). The extracts of the bark and leaves were able to interfere with the initial stages of biofilmformation, but there was no interference of the extract on the established biofilm. There was a high sensitivity of Staphylococcus spp. isolates from subclinical mastitis cases in ruminants when subjected to the extracts from bark and leaves of Commiphora leptophloeos, as well as regarding the ability of extracts to interfere in biofilm formation, indicating their potential in the use for ruminant mastitis therapy.
Keywords: Mastitis, Staphylococcus spp., imburana de cambão, phytotherapy, biofilm.
Recebido em: 14 de fevereiro de 2019.
Aceito em: 17 de junho de 2019.
Introdução
A mastite é uma das doenças infecciosas que mais tem causado prejuízos à indústria de laticínios em todo o mundo, ocasionando perdas profundas em todas as etapas da produção(1, 2, 3). Os efeitos negativos dessa doença decorrem do alto custo de seu tratamento e da administração de antibióticos, resultando também em diminuição da produção de leite, descarte do leite contaminado por agentes antimicrobianos e surgimento de bactérias resistentes(4, 5). A manifestação clínica da mastite é caracterizada por alterações físicas nas glândulas mamárias(6). A forma subclínica não apresenta sintomas e é determinada por uma alta contagem de células somáticas no leite. A mastite subclínica reduz a qualidade do leite e pode disseminar a infecção bacteriana entre diferentes propriedades e espécies(7). O diagnóstico e o controle são complexos e exige desinfecção dos tetos, contagem de células somáticas, análise bacteriológica do leite, antibioticoterapia e abate de vacas com infecção crônica(8, 9, 7). A mastite subclínica tem uma etiologia multifatorial, com infecção bacteriana frequentemente relatada na literatura. Espécies do gênero Staphylococcus são as bactérias mais comumente isoladas em casos de infecção intramamária em ruminantes(10, 11).
O tratamento da mastite é complexo e requer o uso de agentes antimicrobianos de amplo espectro com ação sistêmica e localizada. No entanto, uma das principais repercussões do emprego destes antibióticos no tratamento de doenças infecciosas é a possibilidade de surgimento de cepas resistentes(12, 13). A tolerância a agentes antimicrobianos deve-se aos genes de resistência presentes nos cromossomos ou plasmídeos bacterianos. Estes genes permitem que as bactérias expressem enzimas capazes de destruir moléculas antimicrobianas, modificando os sítios alvo das drogas ou criando uma rota metabólica alternativa(14,15). Os genes mais importantes associados à resistência a Staphylococcus spp. estão relacionados com a produção de β-lactamase, que, em alguns casos, pode ser mediada pelo gene blaZ, conferindo resistência aos agentes antimicrobianos da classe β-lactâmicos e levando à formação de biofilme, um dos principais fatores responsáveis de infecção bacteriana persistente ou crônica(16, 17, 18).
Devido às preocupações atuais em relação ao surgimento de bactérias resistentes a antibióticos comerciais comumente empregados no tratamento de mastite subclínica em ruminantes, a busca por novos compostos com ação antibacteriana e seu potencial para reduzir ou prevenir a formação de biofilme é particularmente importante no contexto da produção animal. De fato, o mercado farmacêutico tem procurado novas fontes de metabólitos com atividade antimicrobiana, como as plantas usadas na medicina popular, que atendem aos mesmos critérios de eficácia, segurança e controle de qualidade que os compostos sintéticos(8, 19, 20, 21).
Commiphora leptophloeos (Mart.) J. B. Gillett é uma árvore espinhosa comumente conhecida como Imburana de Cambão e Imburana de espinhos, pertencentes à família Burseraceae, que compreende mais de 150 espécies(22, 23). Adaptada a regiões tropicais e subtropicais, esta espécie é conhecida por suas propriedades medicinais, relacionadas à presença de compostos bioativos no metabolismo vegetal, tradicionalmente utilizados em comunidades locais no nordeste do Brasil como infusão, chá ou xarope para o tratamento de doenças infecciosas e sintomas inflamatórios(24, 25, 26). Apesar de seu valor terapêutico na medicina popular, ainda existem poucos relatos científicos sobre a atividade farmacológica da Commiphora leptophloeos e seu potencial bioativo, justificando um interesse emergente em estudar o potencial antimicrobiano desta espécie como uma possível alternativa no tratamento da mastite em ruminantes. Portanto, o presente estudo teve como objetivo avaliar a atividade antimicrobiana in vitro de extratos etanólicos da casca e folhas de Commiphora leptophloeos (Mart.) J. B. Gillett frente Staphylococcus spp. isolados de casos de mastite subclínica em ruminantes, bem como realizar uma avaliação fitoquímica da composição do extrato etanólico bruto (EEB) das cascas e folhas de C. leptophloeos.
Material e métodos
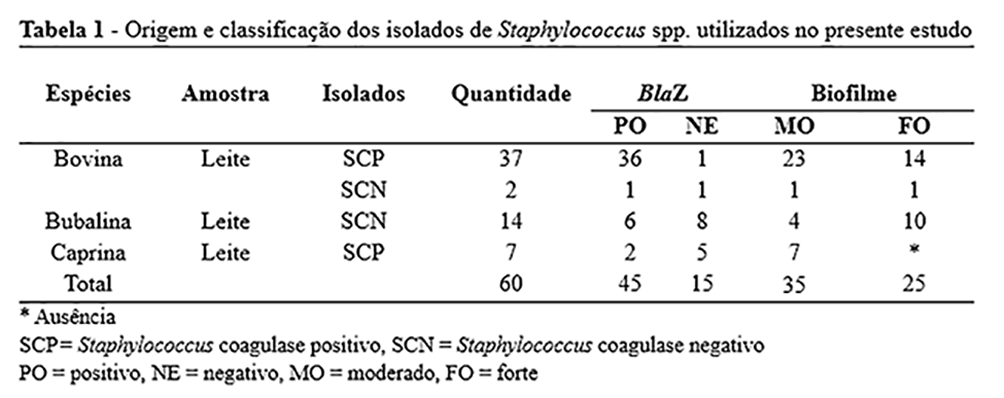
Sessenta isolados de Staphylococcus spp. foram obtidos de casos de mastite subclínica em bovinos (39), búfalos (14) e caprinos (07) em laticínios da região Nordeste do Brasil, sendo armazenados no Laboratório de Microbiologia e Imunologia Animal do Departamento de Medicina Veterinária da Universidade Federal do Vale do São Francisco (UNIVASF), Brasil. Dezesseis isolados foram classificados como Staphylococcus spp. coagulase negativa (SCN) e 44 como coagulase positiva (SCP). Os isolados foram divididos quanto a presença / ausência do gene blaZ, bem como também a uma forte / moderada formação de biofilme (Tabela 1).
O método de Kirby-Bauer foi empregado com base nas recomendações do CLSI(27) para os testes de sensibilidade in vitro em comparação aos agentes comerciais. Ampicilina (10 μg), penicilina G (10 μg) e oxacilina (10 μg) (Laborclin®) foram utilizadas para determinar o perfil de resistência dos isolados aos principais β-lactâmicos rotineiramente utilizados para o tratamento da mastite subclínica. A zona de inibição foi medida e comparada com a tabulação do CLSI(27) para a determinação da resistência ou sensibilidade da amostra ao antimicrobiano. Os resultados foram expressos como resistentes ou sensíveis aos antibióticos, seguindo a recomendação do fabricante do disco.
Os extratos etanólicos brutos foram obtidos da casca e folhas de plantas de C. leptophloeos coletadas no município de Petrolina (PE), no Campus de Ciências Agrárias da UNIVASF, no âmbito do Perímetro de Irrigação C1 (09º 19 '47.22' 'S, 40 ° 33' 22.39 "W, elevação: 392 m), localizada na região semiárida (bioma Caatinga – Figura 1). A casca e as folhas foram secas em estufa de secagem com circulação de ar a 40 °C por cinco dias. As amostras de casca (446 g) e folhas (215 g) foram então trituradas separadamente e submetidas à extração etanólica, seguida de filtração e concentração em evaporador rotativo a uma temperatura média de 50 °C para obtenção dos extratos etanólicos brutos da casca (105,56 g) e folhas (45,99 g). Quatro extrações foram realizadas em intervalos de 72 horas para a extração máxima dos constituintes químicos. Os extratos foram diluídos em etanol a 95% e utilizados nos ensaios de atividade antimicrobiana in vitro.
Para a análise dos dados pelo método HPLC-DAD, o extrato de folhas de C. leptophloeos foi extraído em SPE C-18 (extração em fase sólida). O cartucho foi ativado com 10 ml de metanol e 10 ml de água ultrapura. Aproximadamente 420mg de extrato foram solubilizados em 500µL de água acidificada (pH 2 com HCl) e 500µL de metanol. Após a aplicação do extrato no cartucho, adicionou-se 10 mL de água ultrapura e a fração com os compostos orgânicos foi eluído com 10 mL de metanol. O solvente foi evaporado, obtendo-se 84,8 mg. Esta fração metanólica foi analisada (CLAE-DAD e LC-MS) utilizando um HPLC Shimadzu Prominence (LC-20AT®)(1) com um detector de arranjo de diodos (SPDM20®)(2). A separação cromatográfica foi realizada em uma coluna Luna C-18 (Phenomenex) medindo 250 mm x 4,6 mm x 5 µm, sendo a fase móvel uma mistura de H2O: ácido acético (99: 1, solvente A) e metanol (solvente B). O extrato de folhas (SPE) foi analisado usando LC-MS. LC-ESI-MS / MS foi obtido no modo negativo. O extrato etanólico de casca (200 mg) foi dissolvido em água acidificada (pH 2 com HCl) e submetido à extração em fase sólida (SPE C-18), sendo previamente ativado e eluído com 10, 30, 50, 70 e 100% de acetona e água. O extrato etanólico bruto, bem como as frações de 10% (23,9 mg), 30% (55,1 mg) e 50% (16,0 mg) foram submetidos análise de massas de alta resolução realizada utilizando um espectrômetro de massa MALDI-TOF (Autoflex III, Bruker Daltonics, Billerica, MA, USA®)(3). Os compostos foram identificados por dados MS através de fragmentação e medições de massa precisas utilizando um calibrante interno (mistura de peptídeos pradão).
A microdiluição foi realizada em microplacas utilizando o método descrito no padrão M07-A9 do manual do Clinical and Laboratory Standards Institute(28). Um total de 0,25 g do extrato de casca e folhas foi diluído separadamente em 10 mL de etanol (95%), obtendo-se extratos em estoque na concentração de 25.000 μg/mL. Seguindo a metodologia, os inóculos foram preparados a partir de uma suspensão bacteriana com turbidez equivalente a 0,5 nos padrões de Mc Farland e levados a uma estufa de circulação de ar a 37 °C/24h sob condições aeróbicas. Para a determinação da CBM, alíquotas (10 μl) foram retiradas do material líquido, semeadas em ágar Mueller-Hinton e incubadas novamente (37 °C/24h). A CBM foi definida como a menor concentração do extrato da casca e das folhas com potencial bactericida contra Staphylococcus spp. Os testes foram realizados em triplicata, e um controle de etanol a 95% foi considerado.
A capacidade de o extrato interferir na formação do biofilme foi determinada pelo método descrito por Nostro et al.(29). Os isolados bacterianos foram cultivados em 3mL de caldo de ágar soja Trypticase (TSA) contendo glicose (1%) e incubados a 37 °C por 24 horas. 100 μL da solução foram adicionados aos poços da microplaca que receberam anteriormente 100 μL do meio de cultura (controle negativo). A concentração do extrato foi metade do valor de CBM determinado no ensaio de microdiluição (0,5 CBM). Após 24 horas de incubação a 37 °C, os poços foram lavados três vezes com 200 µL de água destilada, corados com 100 µL de violeta de genciana (0,25%) durante três minutos e lavados novamente com 200 µL de água destilada. Posteriormente, 200 μL de álcool-acetona foram utilizados para diluir os cristais violeta. A absorvância foi medida num leitor de microplacas (kit Elisa Easy®) a 620 nm. S. epidermidis ATCC 12228 foi usado como controle negativo, enquanto S. aureus ATCC 25923 foi usado como controle positivo
A capacidade dos extratos em interferirno biofilme consolidado foi determinada com base no método descrito por Nostro et al.(29). O biofilme foi formado a partir da incubação de 100 μL de inóculo bacteriano em poços de microplacas a 37 °C por 24 horas. Os poços foram lavados três vezes com água destilada para a remoção de células não aderidas e então 200 μL do extrato (0,5 CBM) foram adicionados. A DO foi determinada imediatamente após esse procedimento, assim como 24 horas após a adição do extrato. A absorvância foi medida num leitor de microplacas (modelo Expert Plus, Biochrom®)(4) a 620 nm. O efeito do extrato sobre o biofilme consolidado foi definido pela seguinte equação: DO média dos poços tratados / DO média dos poços de controle x 100.
O teste de normalidade Kolmogorov-Smirnov revelou ausência de normalidade dos dados referentes a CBM dos dois tipos de extrato. A transformação logarítmica de base 10 (log10) foi realizada, mas também foi insuficiente para remover a heterogeneidade das variâncias. Portanto, o teste não-paramétrico de Wilcoxon foi usado para comparações pareadas de valores médios de CBM dos dois extratos. O teste de Mann-Whitney foi utilizado para as comparações da CBM entre os isolados (coagulase-positivo x coagulase negativa; blaZ presente x blaZ ausente). Para análise dos dados, foi utilizado o programa Statistical Package for the Social Sciences (SPSS) versão 20.0 para Windows
Resultados
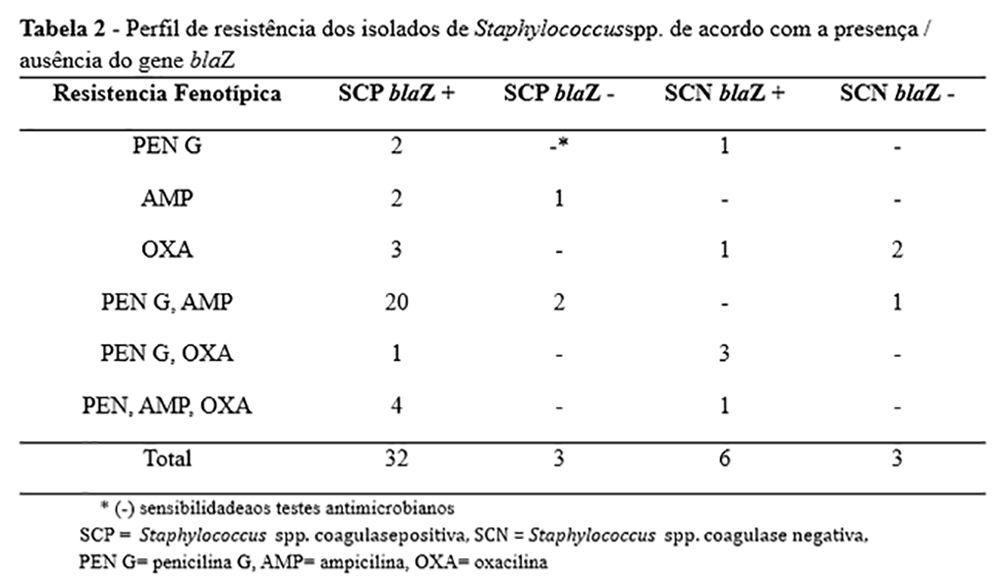
De todos os isolados empregados neste estudo, 34 apresentaram resistência à penicilina, 30 à ampicilina e 15 à oxacilina, e vários isolados foram resistentes a mais de um antimicrobiano (Tabela 2). Os resultados do teste de sensibilidade foram comparados com base na classificação genotípica para a presença do gene blaZ. Entre os 46 isolados com o gene blaZ, apenas dez foram sensíveis a todos os antimicrobianos β-lactâmicos testados (Tabela 2).
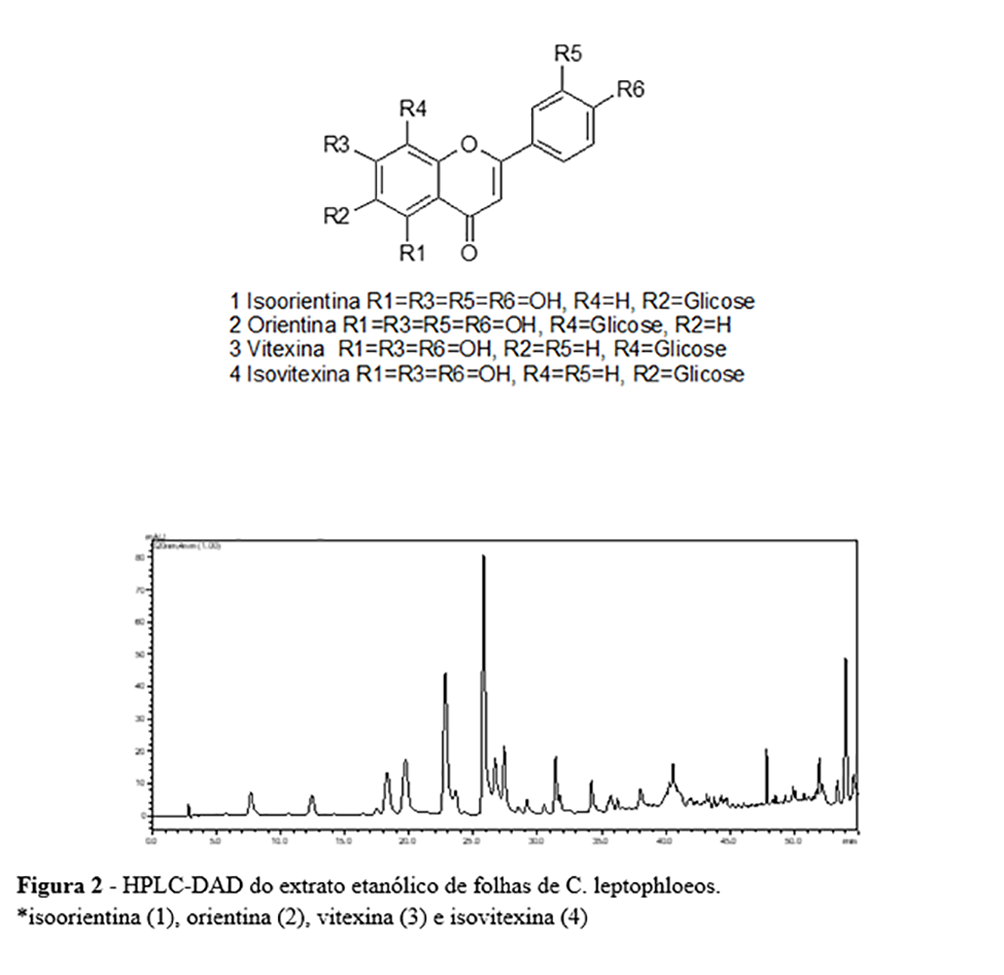
A análise dos dados de HPLC-DAD e HPLC-ESI-MS (Tabela 3) permitiu a identificação dos quatro principais componentes das folhas de C. leptophloeos, que foram as C-glicosil flavonas, isoorentina, orientina, isovitexina e vitexina. O espectro ESI-MS no modo negativo, referindo-se aos Picos 1 e 2 (Figura 2 e Tabela 3), exibiu o mesmo valor no espectro de massa a m/z 447, sugerindo que estes compostos são os isómeros isoorientina e orientina. O mesmo foi encontrado para os picos 3 e 4 a m/z 431, sugerindo que estes são os isômeros vitexina e isovitexina. Outros picos de fenólicos apareceram nos espectros, mas a identificação não foi possível. Os flavonóides foram previamente isolados e identificados nas partes aéreas de C. leptophloeos(30). No entanto, esta é a primeira vez que os flavonóides C-glicosil foram relatados no gênero Commiphora.
Os espectros MALDI-TOF e MALDI-MS / MS permitiram a identificação de unidades de flavan-3-ol e ligações de galoil nos oligômeros. Taninos condensados (proantocianidinas) foram identificados na fração obtida com 10% de acetona, com a ocorrência de unidades de prorobinetinidina (Série A) (Figura 3), devido à perda consecutiva de 288u. Os picos foram observados de duas unidades (m / z 601, [M + Na] +) até sete unidades (m / z 2041 [M + Na] +) de prorobinetinidina. Proantocianidinas da série C também foram encontradas nesta fração (Figura 3), com duas unidades de prorobinetinidina e uma unidade de prodelfinidina (m / z 905 [M + Na] +).
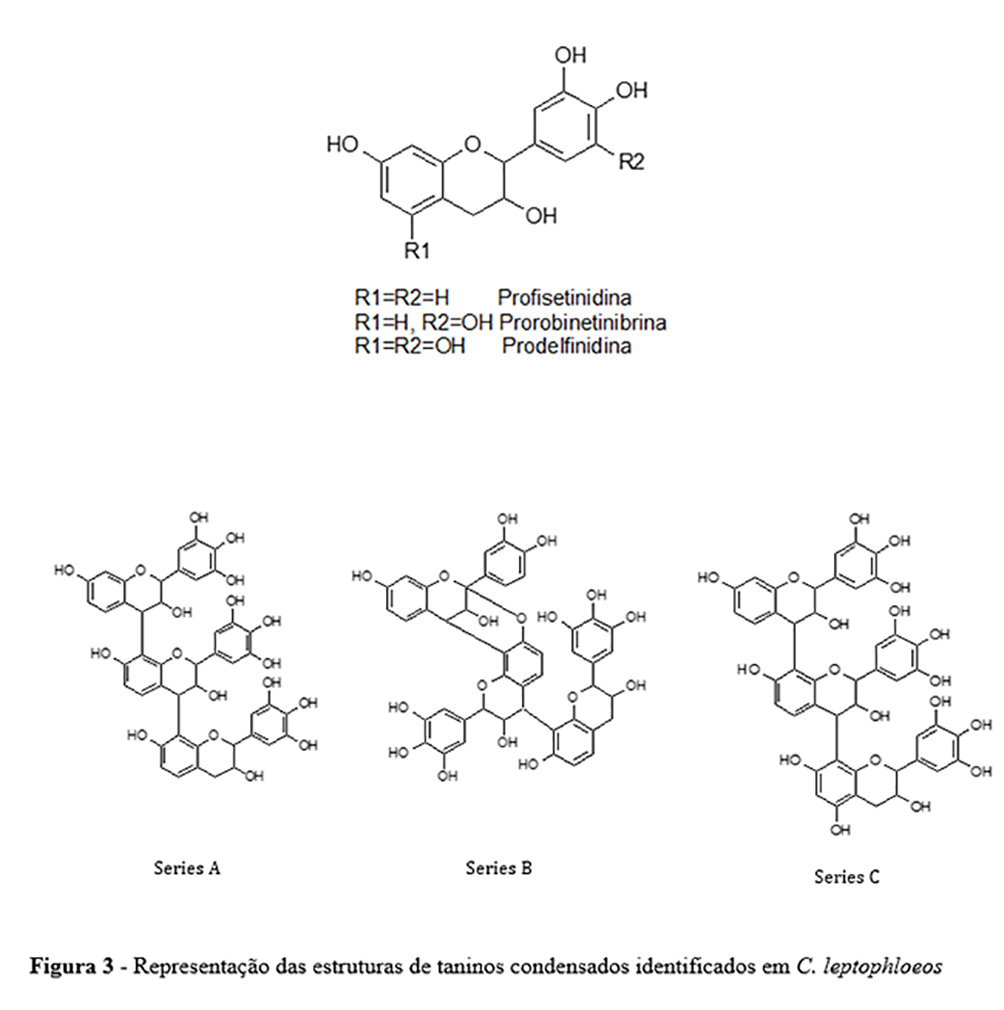
Para esta série, os picos foram encontrados em m / z 1193 [M + Na] +, correspondendo a três unidades de prorobinetinidina e uma de prodelfinidina; m / z 1481 [M + Na] +, quatro unidades de prorobinetinidina e uma de prodelfinidina; m/z 1769 [M + Na] +, cinco unidades de prorobinetinidina e uma de prodelfinidina; e m/z 2057 [M + Na] +, seis unidades de prorobinetinidina e uma de prodelfinidina. A fração obtida com 50% de acetona exibiu apenas um pico a m/z 871 [M + Na] +, correspondendo a uma unidade de profisetinidina e duas unidades de prorobinetinidina. A figura 3 mostra as estruturas poliméricas, que foram confirmadas pelos espectros MS / MS. A fração obtida com 50% de acetona apresentou perfil com taninos hidrolisáveis, diferindo das demais frações analisadas. Os espectros MS / MS demonstraram picos com perdas de 152µ, o que corresponde ao ácido gálico e confirma a presença de galotaninos na fração. Os dados atuais sobre taninos condensados estão de acordo com os achados relatados anteriormente para a casca de C. leptophloeos(31). No entanto, a presença de galotaninos na casca de C. leptophloeos é relatada pela primeira vez na presente investigação.
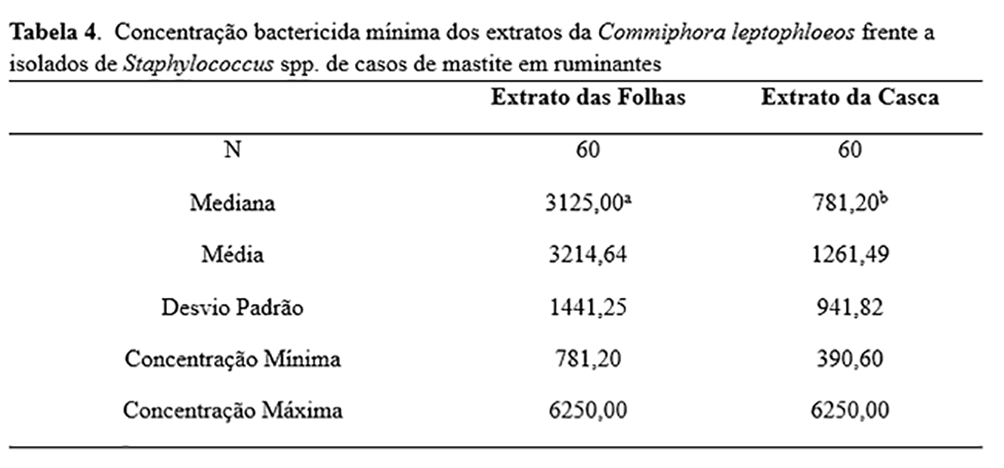
A diferença na CBM entre os dois extratos foi estatisticamente significante (p <0,001), com valores mais baixos sendo encontrados para o extrato de casca (Tabela 4). Ambos os extratos demonstraram ação antimicrobiana contra todos os isolados de Staphylococcus spp. A melhor atividade do extrato das folhas ocorreu na concentração de 3.125 μg/mL (36/60), e a melhor atividade do extrato de casca foi obtida na concentração de 781.2 μg/mL (24/60).
Nenhuma diferença estatisticamente significativa foi encontrada nos valores médios da CBM para os isolados SCP e SCN (1207,00 e 1539,00 μg/mL, respectivamente) usando o extrato da casca. No entanto, uma diferença significativa foi encontrada para o extrato das folhas (p <0,05), com médias de 2823,00 e 3814,00μg/mL para SCP e SCN, respectivamente. Uma diferença estatisticamente significativa foi encontrada para o extrato da casca em relação aos isolados com e sem o gene blaZ (p <0,05), pois os valores médios de CBM foram 1146,00 e 1674,00μg/mL, respectivamente.
O extrato das folhas foi capaz de diminuir a formação de biofilme entre 43 isolados, 20 dos quais foram classificados como produtores fortes de biofilme e 20 classificados como produtores moderados. O extrato da casca foi capaz de interferir na formação do biofilme em 47 isolados, 22 dos quais classificados como produtores fortes e 25 classificados como produtores moderados (Tabela 1). Nenhum extrato exerceu efeito sobre o biofilme consolidado.
Discussão
Dez dos 46 isolados positivos para a presença do gene blaZ foram sensíveis a todos os agentes antimicrobianos testados. Esse achado pode ser justificado pela ausência do complexo completo blaZ-blaR1-blaI de genes reguladores(32). A β-lactamase codificada pelo gene blaZ é regulada na presença de um antibiótico, através dos genes adjacentes blaI, um repressor da transcrição blaZ e o antirrepressor blaR1.Outro ponto importante na resistência do Staphylococcus spp., a outros antimicrobianos pode estar relacionado ao seu uso indiscriminado, com a frequência e o emprego a longo prazo desses fármacos no tratamento de infecções intramamárias promovendo a seleção de cepas resistentes devido à produção de β-lactamase codificada pelo gene blaZ(33, 34).
No presente estudo, os extratos das folhas e da casca foram capazes, respectivamente, de reduzir a formação de biofilme em 43 e 47 isolados classificados de moderados a fortes produtores de biofilme. Como o extrato bruto etanólico das folhas exibia quatro flavonóides principais, pode-se especular que essa atividade poderia ser atribuída a esses compostos, como Awololaet al.(35) demonstraram a atividade de formação de antibiofilme de isovitexina (um flavonóide primário encontrado na casca de C. leptophloeos no presente estudo) para S. aureus ATCC 29213. Trentin et al.(36) relatam a interferência do extrato aquoso da casca de C. leptophloeos na formação de biofilme de isolados de Staphylococcus epidermidis. Em outro estudo, Trentin et al.(31) também verificaram a atividade de formação de antibiofilme do extrato aquoso da casca de C. leptophloeos aplicado a isolados de Pseudomonas aeruginosa, e os autores também relataram a presença de taninos condensados no extrato, o que está de acordo com os presentes achados. Além disso, a presença de galotaninos na casca de C. leptophloeos é relatada pela primeira vez na presente investigação. As plantas têm sido cada vez mais pesquisadas com o objetivo de inibir a formação do biofilme, por se mostrarem potencialmente eficazes no que se refere à sua atividade antimicrobiana sobre várias espécies de microrganismos(37).
Os extratos etanólicos brutos da casca e folhas de C. leptophloeos demonstraram atividade contra todos os isolados de Staphylococcus testados, com diferença significativa em termos do CBM, que foi menor para o extrato de casca (781,2 μg/mL). Este achado deve-se ao fato de que a casca de C. leptophloeos contém taninos condensados (proantocianidinas) que são hidrolisáveis. Entre os isolados blaZ positivos, pode-se observar uma acentuada atividade antimicrobiana, quando utilizou-se o extrato das cascas. Assim, os resultados demonstram que extratos de C. leptophloeos são potenciais agentes antimicrobianos. No entanto, mais estudos são necessários para confirmar a eficácia destes extratos contra isolados blaZ positivos. No estudo realizado por Trentin et al.(36), o extrato aquoso da casca de C. leptophloeos apresentou atividade antimicrobiana contra S. epidermidis na concentração de 1,0 mg/mL. As diferenças na CBM podem ser atribuídas à composição química das substâncias antimicrobianas em diferentes extratos vegetais, mesmo quando da mesma espécie. Uma análise de compostos fenólicos realizada por Araujo et al.(38) revelou alto teor de flavonóides e taninos na casca de C. leptophloeos. Os compostos fenólicos presentes nas plantas estão relacionados à proteção e oferecem alta resistência a microrganismos e pragas(39).
Outras plantas nativas do bioma Caatinga também foram testadas e todas demonstraram resultados antimicrobianos positivos. Sá et al.(40) avaliaram a atividade antimicrobiana de seis espécies (Amburana cearensis, Neoglazio viavariegata, Encholirium spectabile, Bromelia laciniosa, Hymenaea martiana e Selaginella convolute) frente a bactérias Gram-positivas e Gram-negativas, em que a A. cearensis e N. variegate apresentaram os menores valores de CBM.
No presente estudo, os Staphylococcus spp. coagulase positivos e os isolados blaZ positivos foram mais sensíveis ao extrato de casca de C. leptophloeos. Esse achado é importante quando se considera o papel do gênero Staphylococcus na etiologia da mastite subclínica em ruminantes(41, 42).O surgimento de cepas resistentes à terapia antimicrobiana convencional ressalta a importância de buscar outras alternativas, como os produtos naturais(43), como observado no presente estudo.
Uma das principais causas da persistência de Staphylococcus spp. nas infecções intramamárias é a sua capacidade de formar biofilme, o que torna essas bactérias resistentes aos agentes antimicrobianos usados no tratamento da mastite(44, 45). Descobriu-se que o biofilme é mais resistente a antimicrobianos quando comparado as células planctônicas(46). Portanto, há uma necessidade urgente de novos agentes antibiofilme(47). No entanto, a forte adesão das bactérias às superfícies dificulta a remoção do biofilme consolidado, o que pode explicar por que os extratos aqui testados não tiveram efeito sobre o biofilme consolidado. Uma vez que quando ocorre à consolidação do biofilme torna-se muito difícil tratar clinicamente os animais, pois as bactérias estão protegidas contra a resposta imune do hospedeiro, bem como dos agentes antimicrobianos(48, 49). Segundo Gonçalez-Ortiz et al.(50), a inibição da formação de biofilme depende de fenômenos ligados à inibição enzimática de proteases e mecanismos de quorum sensing. A estabilidade do biofilme é devida à deposição de proteínas e carboidratos, o que dificulta a ação de compostos sanificantes e antimicrobianos(51).
Conclusão
Com base nos resultados encontrados, os extratos etanólicos da casca e folhas de C. leptophloeos apresentaram ação antimicrobiana contra cepas de Staphylococcus spp., bem como capacidade de inibir a formação de biofilme. Portanto, é necessário realizar estudos adicionais de toxicidade sobre a ação antimicrobiana desta espécie, tanto in vitro quanto in vivo, a fim de avançar na investigação fitoquímica das frações do extrato utilizando diferentes solventes, além de técnicas mais avançadas, como biomarcadores, destacando a extração de aminoácidos e proteínas, caracterizando melhor o potencial dessa planta contra agentes causadores de mastite em ruminantes, com o objetivo de desenvolver produtos que possam ser utilizados para o tratamento de animais afetados.
Agradecimentos
Os autores agradecem aos seguintes órgãos brasileiros de fomento: Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão de bolsa de mestrado, ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), a Fundação de Amparo à Ciência e Tecnologia de Pernambuco (FACEPE) e o Centro de Tecnologias Estratégicas do Nordeste (CETENE).
Conflito de interesses
Este manuscrito nunca foi publicado. Os autores estão de acordo com as declarações e requisitos para publicação. Nenhum dos autores tem qualquer conflito de interesse a declarar.
Referências
1 Kościuczuk EM, Lisowski P, Jarczak J, Krzyżewski J, Zwierzchowski L, Bagnicka E. Expression patterns of β-defensin and cathelicidin genes in parenchyma of bovine mammary gland infected with coagulase-positive or coagulase-negative Staphylococci. BMC Veterinary Research.2014;10:246.
2 Nagahata H, Kawai H, Higuchi H, Kawai K, Yayou K, Chang CJ. Altered Leukocyte Responsiveness in Dairy Cows with Naturally Occurring Chronic Staphylococcus aureus Mastitis. Journal of Veterinary Medical Science. 2011;73:885-94.
3 Ruiz AK, Peña J, González D, Ponce P. Prevalence, somatic cell count and etiology of bovine mastitis in Cuban herds from Mayabeque province using hand and machine milking. Revista Salud. Animal. 2014;36:7-13.
4 Dhakal IP, Dhakal P, Koshihar T; Nagahata H. Epidemiological and bacteriological survey of buffalo mastitis in Nepal. Journal Veterinary Medical Science. 2007; 69:1241–1245.
5 Gernand E, Rehbein P, Von Borstel UU, König S. Incidences of and genetic parameters for mastitis, claw disorders, and common health traits recorded in dairy cattle contract herds. Journal of Dairy Science. 2012; 95:2144–56.
6 Falentin H, Rault L, Nicolas A, Bouchard DS, Lassalas J, Lambe P, et al. Bovine Teat Análise Microbiome revelaram redução da Diversidade Alpha e mudanças significativas nos perfis taxonômicos em Quarters com uma história de mastite. Frontiers in Microbiology. 2016; 7:480.
7 Reyes-Jara A, Cordero N, Aguirre J, Troncoso M, Figueroa G. Antibacterial Effect of Copper on Microorganisms Isolated from Bovine Mastitis. Frontiers Microbiolgy. 2016;7:626.
8 Gomes F, Henriques M. Controle da mastite bovina: Abordagens terapêuticas antigas e recentes. Current Microbiology. 2016; 72:377-382.
9 Nickerson SC. Control of heifer mastitis: antimicrobial treatment-an overview. Veterinary Microbiology. 2009;134:128–135.
10 Godden SM, Jansen JT, Leslie KE, Smart NL, Kelton DF. The effect of sampling time and sample handling on the detection of Staphylococcus aureus in milk from quarters with subclinical mastitis. Canadian Veterinary Journal. 2002; 43:38-42.
11 Tenhagen BA, Köster G, Wallmann J, Heuwiese RW. Prevalence of mastitis pathogens and their resistance against antimicrobial agents in dairy cows in Brandenburg, Germany. Journal Dairy Science. 2006;89:2542-2551.
12 Goni P,Vergara Y, Ruiz J, Albizu I, Vila J, Gomez-Lus R. Antibiotic resistance and epidemiological typing of Staphylococcus aureus strains from ovine and rabbit mastitis. International Journal Antimicrobial Agents. 2004; 23:268-272.
13 Harbottle H, Thakura S, Zhao S, White DG. Genetics of antimicrobial resistance. Animal Biotechnology. 2006;17:111-124.
14 Sheldon AT, Antibiotic resistance: a survival strategy. Clinical Laboratory Science. 2005;18: 170-180.
15Tenover F.C. & McGowan JR.J.E. 2008. Antimicrobial resistance. International Encyclopedia of Public Health. 2008; p.211-219.
16 Olsen JE, Christensen H, Aarestrup FM. Diversity and evolution of blaZ from Staphylococcus aureus and coagulase negative staphylococci. Journal Antimicrobial Chemotherapy. 2006;57:450–460.
17 Wright GD. Bacterial resistance to antibiotics: enzymatic degradation and modification. Advanced Drug Delivery Reviews. 2005;57:1451-1470.
18 Xue T, Chen M, Shang F. Short communication: effects of lactose and milk on the expression. of biofilm-associated genes in Staphylococcus aureus strains isolated from a dairy cow with mastites. Journal Dairy Science. 2014;97:6129-6134.
19 Mendes LPM, Maciel KM, Vieira ABR, Mendonça LCV, Silva RMF, Rolim Neto PJ, et al. Atividade Antimicrobiana de Extratos Etanólicos de Peperomia pellucida e Portulaca pilosa. Revista Ciências Farmacêuticas Básica. 2011;32:121-125.
20 Mordmuang A, Shankar S, Chethanond U, Voravuthikunchai SP. Effects of Rhodomyrtus tomentosa Leaf Extract on Staphylococcal Adhesion and Invasion in Bovine Udder Epidermal Tissue Model. Nutrients. 2015;7:8503–8517.
21 Xu W, Guan R, Lu Y, Su X, Xu Y, Du A, Hu S. Therapeutic effect of polysaccharide fraction of Atractylodis macrocephalae Koidz. in bovine subclinical mastitis. BMC Veterinary Research. 2015;1:165.
22 Maia GN. Caatinga: árvores e arbustos e suas utilidades. São Paulo: Leitura & Arte, 2004; 413p.
23 Sampaio EVSB Pareyn FGC, Figueirôa JM, Santos Junior AG. Espécies da flora nordestina de importância econômica potencial. Recife. Associação Plantas do Nordeste, 2005; 331p.
24Agra MF, Baracho GS, Silva NK, Basílio IJLD, Coelho VPM. Medicinais e venenosas diversidades da flora de "Cariri Paraibano", Brasil. Journal of Ethnopharmacology.2007; 111: 383-395.
25Abdel-Daim MM, Ghazy EW, Fayez M. S. Synergistic protective role of mirazid (Commiphora molmol) and ascorbic acid against tilmicosin-induced cardiotoxicity in mice. Canadian Journal of Physiology and Pharmacology. 2015; 93: 45-51.
26Souza Pereira JJ, Pereira APC, Jandus, JJB, da Paz JA, Crovella S, Correia MTS, Silva JA. Commiphora leptophloeos Phytochemical and Antimicrobial Characterization. Frontiers in Microiology. 2017; 8: 1-10.
27 Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Ninth Edition. CLSI document M07-A9. Wayne, PA: Clinical and Laboratory Standards Institute.2012; 88p.
28 Clinical and Laboratory Standards Institute (CLSI). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; Approved standards. CLSI Document M7-A7. Wayne, PA Pennsylvania. 2006; 88p.
29Nostro A, Roccaro AS, Bisignano G, Marino A, Cannatelli MA, Pizzimenti FC, Cioni, PL, Procopio, F, Blanco, AR. Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of Medical Microbiology. 2007; 56:519–523.
30 Souza MP, Machado MIL, Braz-Filho R. Six flavonoids from Bursera leptophloeos. Phytochemistry. 1989,28: 2467-2470.
31 Trentin DS, Silva DB, Amaral MW, Zimmer KR, Silva MV, Lopes NP, et al. Taninos possuindo bacteriostatic Efeito Impair Pseudomonas aeruginosa adesão e formação de biofilme. Plos One. 2013;8:66-257.
32Mendonça ECL, Marques VF, Melo DA, Alencar TA, Coelho IS, Coelho SMO, Souza MMS. Caracterização fenogenotípica da resistência antimicrobiana em Staphylococcus spp. isolados de mastite bovina. Pesquisa Veterinária Brasileira.2012;32:859-864.
33Pitkälä A, Haveri M, Pyörälä S, Myllys V, Honkanen-Buzalski, T. Bovine mastitis in Finland 2001—Prevalence, distribution of bacteria, and antimicrobial resistance. Journal Dairy Science.2004;87:2433–2441
34Kumar R., Yadav B.R. & Singh R.S. Genetic determinants of antibiotic resistance in Staphylococcus aureus isolates from milk of mastitic crossbred cattle. Current Microbioloy. 2009;60:379–386.
35 Awolola GV, Koorbanally NA, Chenia H, Shode FO, Baijnath H. Antibacterial and Anti-Biofilm Activity of Flavonoids and Triterpenes Isolated from the Extracts of Ficus sansibarica Warb. Subsp. Sansibarica (Moraceae) Extracts. African Journal of Traditional, Complementary, and Alternative Medicines. 2014;11:124–131
36 Trentin DS, Giordani RB, Zimmer KR, Silva AG, Silva MV, Correia MT, et al. Potential of medicinal plants from the Brazilian semi-arid region (Caatinga) against Staphylococcus epidermidis planktonic and biofilm lifestyles. Journal of Ethnopharmacology. 2011;137:327-335.
37 Pereira JV, Pereira MSV, Sampaio FC, Sampaio MCC, Alves PM, Araújo CRF, Higino JS. Efeito antibacteriano e antiaderente in vitro do extrato da Punica granatum Linn. sobre microrganismos do biofilme dental. Revista Brasileira Farmacognosa. 2006;16: 88-93.
38 Araujo TAS, Alencar NL, Amorim ELC, Albuquerque UP. A new approach to study medicinal plants with tannins and flavonoids contents from the local knowledge. Journal of Ethnopharmacology.2008;120:72-80.
39 Everette JD, Bryant QM, Green AM, Abbey YA, Wangila G W, Walker RB. Thorough study of reactivity of various compound classes toward the Folin-Ciocalteou reagent. Journal of Agricultural Food Chemistry. 2010; 58:139-144.
40 Sá MCA, Peixoto RM, Krewer CC, Almeida JRG, Vargas AC, Costa MM. Antimicrobial activity of caatinga biome ethanolic plant extracts against gram negative and positive bacteria. Revista Brasileira de Ciência Veterinária. 2011;18:62-66.
41 Coelho SMO, Reinoso E, Pereira IA, Soares LC, Demo M, Bogni C, Souza MMS. Virulence factors and antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis in Rio de Janeiro. Pesquisa Veterinária Brasileira. 2009; 29:369-374.
42 Taponen S, Pyörälä S. Coagulase-negative staphylococci as cause of bovine mastitis not so different from Staphylococcus aureus? Veterinary Microbiology. 2009;134:29-36.
43 Godreuil S, Leban N, Padilla A, Hamel R, Luplertlop N, Chauffour A, et al. Structure and Antimicrobial Activity against Multidrug Resistant Bacterial Strains. PLoS ONE. 2014; 9:1371.
44 Nasr RA, AbuShady HM, Hussein HS. Biofilm formation and presence of icaAD gene in clinical isolates of staphylococci. Egyptian Journal of Medical Human Genetics. 2012;13:269-274.
45 Terki IK, Hassaine H, Oufrid S, Bellifa S, Mhamed I, Lachachi M,Timinouni M, Detection of icaA and icaD genes and biofilmformation in Staphylococcus spp. isolated from urinary catheters at the University Hospital of Tlemcen (Algeria). African Journal Microbiology Research. 2013;7:5350-5357.
46 Jadhav S, Shah R, Bhave M, Palombo EA. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Journal Food Control. 2013;29:125-130.
47 Schilaci D, Arizza V, Dayton T, Camarda L, Di Stefano V. In vitro anti-biofilm activity of Boswellia spp. oleogum resin essential oils. Letters in Applied Microbiolog. 2008;47:433-8.
48 Drenkard E. Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes and Infection. 2003; 5:1213–1219.
49 Hoyle BD, Costerton JW. Bacterial resistance to antibiotics: the role of biofilms. Progress in Drug Research. 1991;37:91–105.
50 Gonzalez-Ortiz G, Perez JF, Hermes RG, Molist F, Jimenez-Diaz R, Martin-Orue SM. O rastreio da capacidade de ingredientes de alimentos naturais para interferir com a aderência de enterotoxigenic Escherichia coli (ETEC) K88 ao muco intestinal suína. British Journal of Nutrition. 2014; 111:633-642.
51 Wang H, Ding S, Wang G, Xu X, Zhou G. In situ characterization and analysis of Salmonella biofilm formation under meat processing environments using a combined microscopic and spectroscopic approach. International Journal Food Microbiology. 2013;167:293-302.