
Abstract
Muscular dystrophies are hereditary diseases that lead to progressive degeneration of the skeletal musculature. Golden Retriever dogs are used as animal models because they show a hereditary muscle disease similar to muscular dystrophy in humans. Aims: To evaluate the immunostaining of M1 (CD68) and M2 (CD163) macrophages, MHC I, MHC II and, utrophin in muscles of Golden Retriever dogs affected by muscular dystrophy (GRMD). Methods: Samples from 17 male dogs affected by GRMD were divided into GI - dystrophic dogs up to one year of age; and GII - dystrophic dogs over one-year-old. Results: Immunostaining of CD163 was higher than CD68 in both GI and GII. CD68 showed no variation between groups of dystrophic animals. MHC class I immunostaining was most evident in the biceps femoris and triceps brachialis. MHC class II was expressed mildly in four dystrophic muscle types in GI and GII. Utrophin immunostaining was higher in GII. Conclusion: M2 macrophages were one of the main mononuclear inflammatory cells found in dystrophic muscles. The number of M2 in muscles of dogs with GRMD increases with age, linking this cell subtype to permanent muscle damage.
Keywords: Golden Retriever Muscular Dystrophy (GRMD); immunohistochemistry; inflammation; myopathy; muscle.
Resumo
As distrofias musculares são doenças hereditárias que levam à degeneração progressiva da musculatura esquelética. Cães Golden Retriever são usados como modelos animais, pois desenvolvem uma doença muscular hereditária semelhante à distrofia muscular em humanos. Objetivos: Avaliar a imunomarcação dos macrófagos M1 (CD68) e M2 (CD163), MHC I, MHC II e utrofina nos músculos de cães Golden Retriever afetados pela distrofia muscular (DMGR). Métodos: Amostras de 17 cães machos afetados por DMGR foram divididas em cães distróficos GI - até um ano de idade; e GII - cães distróficos com mais de um ano de idade. Resultados: a imunomarcação de CD163 foi maior que CD68 nos grupos GI e GII. CD68 não mostrou variação entre os grupos de animais distróficos. A imunomarcação de MHC classe I foi mais evidente no bíceps femoral e tríceps braquial. O MHC classe II foi expresso discretamente nos quatro tipos de músculo distrófico no GI e GII. A imunomarcação de utrofina foi maior no GII. Conclusão: Os macrófagos M2 foram uma das principais células inflamatórias mononucleares encontradas nos músculos distróficos. O número de M2 nos músculos de cães com DMGR aumentou com a idade, ligando esse subtipo de célula a danos musculares permanentes.
Palavras-chave: Distrofia Muscular do Golden Retriever (DMGR); imuno-histoquímica; inflamação; miopatia; músculo.
Section: Medicina Veterinária
Received
April 4, 2019.
Accepted
April 20, 2020.
Published
June 16, 2020.
www.revistas.ufg.br/vet
visit the website to get the how to cite in the article page.
Introduction
Duchenne muscular dystrophy (DMD) is a severe X-linked genetic disease that leads to degeneration and gradual loss of function of skeletal and cardiac muscles(1). Dystrophin is a protein that links the cytoskeleton to the extracellular matrix. Reduced dystrophin synthesis in affected people leads to progressive muscle weakness(1).
Golden Retriever Muscular Dystrophy (GRMD) was recognized in the 1970s and 1980s. GRMD has phenotypic characteristics similar to those in people affected by Duchenne Muscular Dystrophy (DMD)(2). GRMD dogs have the corresponding spontaneous mutation in the same gene as human patients(2).
Affected dogs show muscle lesions similar to those seen in man, making them a suitable experimental model for human patients(3). Muscle lesions vary depending on the muscle group and stage of the disease in both dogs and people(4). The features of disease in dogs, including elevated serum enzymes, sarcolemma defects and complex repetitive discharges on electromyography correspond with the pathogenesis of disease in human DMD patients(2). In general, cardiorespiratory failure and progressive worsening of symptoms are the cause of death in both dogs and people(5).
The GRMD model has played an important role in the characterization of pharmacological interactions, cellular and immunological reactions, and genetic therapies(2).
Utrophin is a muscle protein that interacts with the same cytoskeletal proteins (dystrophin) but binds to different sites on the actin filament(6). This suggests that utrophin and dystrophin may act similarly inside the cell. In healthy animals, utrophin can be expressed at the neuromuscular junction (NMJ), tendon muscle, in vessels and nerves, as well as participating in muscle formation and regeneration(6, 7). In mdx mice, utrophin compensates for the lack of dystrophin, maintaining the interaction between the actin cytoskeleton and the dystrophin-glycoprotein complex on the sarcolemma. Positive anti-utrophin antibody immunostaining shows variable intensity in dystrophic skeletal muscles(6).
After muscle injury macrophages can promote injury or repair processes, according to their phenotype(8, 9, 10). During muscle regeneration, two types of macrophages (M1 and M2) are involved in myogenesis and angiogenesis. These cells play different roles in muscle repair according to their activation state(11). M1 macrophages (present in the acute phase of muscle injury in mdx mice) are responsible for lysis of myofibers. This occurs by the cytotoxic action of nitric oxide (NO), produced by the transformation of the enzyme induced nitric oxide synthase (iNOS) in the presence of arginine. In later stages of disease, the macrophage population observed is the M2 lineage, which promotes muscle growth and regeneration through competition with M1 for the use of arginine(8, 9). During this phase, M2 macrophages are responsible for phagocytosis and proteolysis of the necrotic material, releasing growth factors which stimulate the migration and activation of satellite cells(12).
The changes in macrophage phenotype in muscular dystrophy demonstrates the importance of these cells in regulating muscle regeneration by direct effects on muscle cells(13). The change in phenotype of the macrophage population from M1 to M2 occurs as the proliferative phase is replaced with the differentiation phase in myogenesis. Thus, the alteration in the phenotype coincides with the regenerative phase and the M2 macrophages are associated with muscle fibers in regeneration. The regulation of this process has previously been attributed to M2 macrophages(8).
The immunophenotypic profile may help our understanding of the pathogenesis of several diseases. Current evidence suggests that the immune response to muscle injury is a complex process, in which several populations of inflammatory cells regulate one another's functions, including muscle regeneration in dogs affected by GRMD. Thus, the transition in the phenotype of macrophages is an important element in in vivo muscle regeneration after acute or chronic injury(12).
Another important element of immune system performance is the major histocompatibility complex (MHC), which is expressed on the surface of mammalian nucleated cells. MHC contains membrane-specific proteins and is important for T lymphocyte activation(14). The MHC gene is divided into class I (MHC I) and class II (MHC II) and these can be identified in cultures and cell suspensions, tissue sections and homogenized tissue immunoblots using specific monoclonal antibodies(15).
During injury large quantities of cytosolic proteins are released into the extracellular space in the dystrophic muscle. These are linked to MHC class I, transforming the myofiber into an antigen-presenting cell(16). The MHCI and MHCII complexes are not expressed in healthy muscle fibers(17).
To gain more insight into the molecular mechanisms responsible for inflammation in muscular dystrophy, we evaluated the immunohistochemical aspects of M1 and M2 macrophages, MHCI, MHCII and utrophin expression in Golden Retriever dogs affected by progressive muscular dystrophy.
Material and Methods
Methodology was in agreement with the ethical principles for animal experimentation adopted by the 'Sociedade Brasileira de Ciência em Animais de Laboratório e Comitê de Ética em Uso Animal' (CEUA) of the Universidade Estadual Paulista "Julio de Mesquita Filho" – Faculdade de Ciências Agrárias e Veterinárias (UNESP/FCAV), number of protocol 025486/09.
The animals were from the 'Associação de Amigos dos Portadores de Distrofia Muscular – AADM' located at the Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo (FMVZ/USP), Ribeirão Preto, Brazil. The samples were classified as dystrophic based on the analysis of genomic DNA held in the 'Centro de Estudos do Genoma Humano' of the Instituto de Biologia, Universidade de São Paulo, Brazil.
Based on previous research, the four muscle groups most frequently affected by muscular dystrophy (masseter, diaphragm, triceps brachialis, and biceps femoris) were analyzed(5).
For histopathologic analysis, muscle samples were fixed in 10% formalin solution, buffered with phosphate (pH 7.4) for 24 hours. After fixation, they were dehydrated in solutions of decreasing concentration of alcohol, diaphanized in xylol and set in paraffin blocks. After histopathologic processing, the muscle samples were stained with Haematoxylin-Eosin (HE).
In canine dystrophy there is hypertrophy of the muscles of the thoracic limbs, tongue, diaphragm, sartorius, and esophagus, and atrophy of the other skeletal muscles(18). In puppies, the flexor muscles are severely affected due to their greater use during this phase of life(19). For this reason, muscle samples from the masseter, diaphragm, triceps brachialis and biceps femoris from 17 male dogs affected by GRMD, were categorized as group I (GI): dystrophic animals up to one year of age (n = 9); and group II (GII): dystrophic animals over one year of age (n = 8).
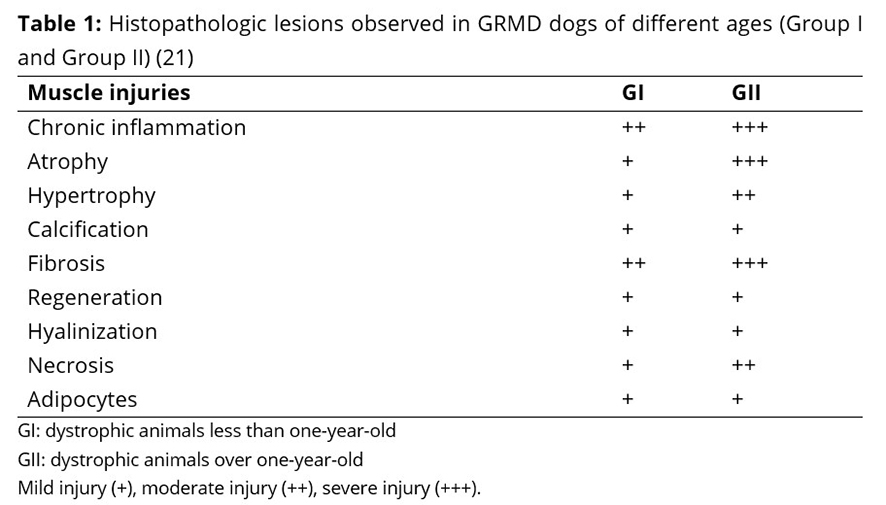
The histopathologic lesions in the animals used in this study have previously been characterized, described and discussed. All the degenerative lesions typical of canine muscular dystrophies were identified and classified by morphological criteria(20, 21). The lesions included hypertrophy, atrophy, skeletal muscle fibrosis, myofiber degeneration, necrosis, mononuclear inflammatory infiltrate, calcification, and hyalinization. The lesions ranged in severity and distribution in the four evaluated muscles, as shown in Table 1(21).
There was no significant difference (P <0.05) in histopathologic lesions in the masseter, diaphragm, triceps brachialis and femoral biceps muscles of dogs affected by muscular dystrophy, indicating that age does not affect the severity of histopathologic lesions described in GRMD dogs(21).
Control muscle samples were obtained from three non-dystrophic Golden Retriever mixed breed dogs, that died from natural causes. Additionally, these dogs were free from any neuromuscular abnormalities.
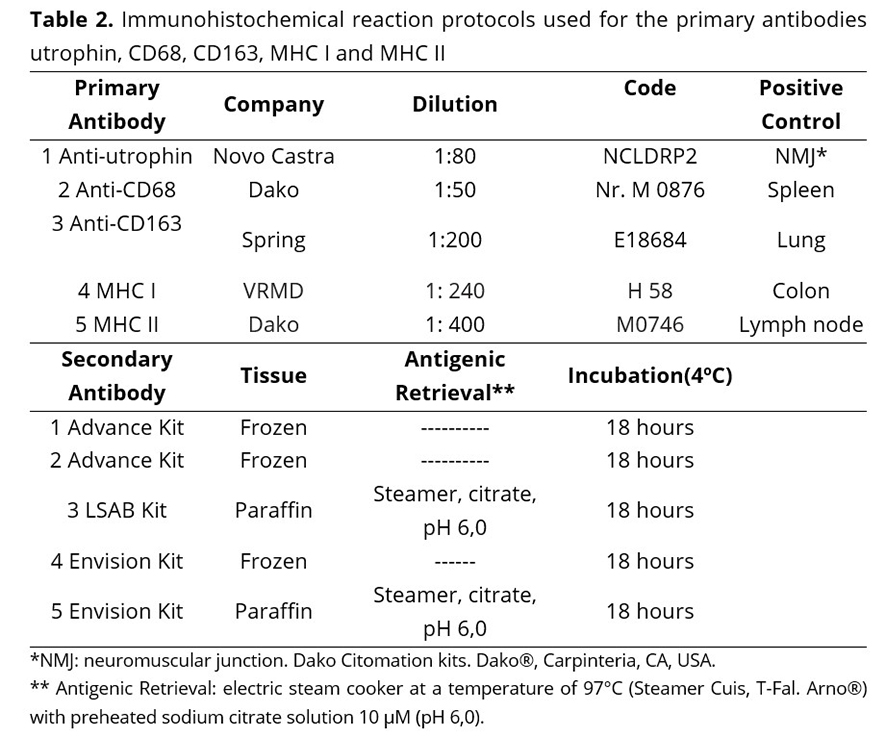
The immunohistochemical method used was the streptoavidin-biotin-peroxidase complex(22). Five primary antibodies were used to evaluate immunostaining of M1 macrophages (CD68), M2 macrophages (CD163), MHC I, MHC II, utrophin, as shown in Table 2.
Positive cells were identified by distinct brown staining in the cytoplasm and membrane. Immunostaining of anti-utrophin, anti-CD68, anti-CD163, MHC I and MHC II, was characterized by the distribution pattern of reaction in the muscle sections. Three immunostaining scores were established according to the intensity and distribution of immunostained cells in muscle fragments(6). For the four muscles evaluated and in all dystrophic dogs, all immunostained cells in ten randomly chosen microscopic fields were counted under light microscopy.
The results from the immunostained cells were submitted to statistical analysis of variance and comparison of means by Tukey test (P <0.05). The means of immunostaining were compared between groups (GI, GII, and control) for each evaluated muscle. For the comparison of means by Tukey test a 5% probability was set. The statistical analyses were performed using SAS software (SAS 9.1, SAS Institute, Cary, NC, USA).
Results
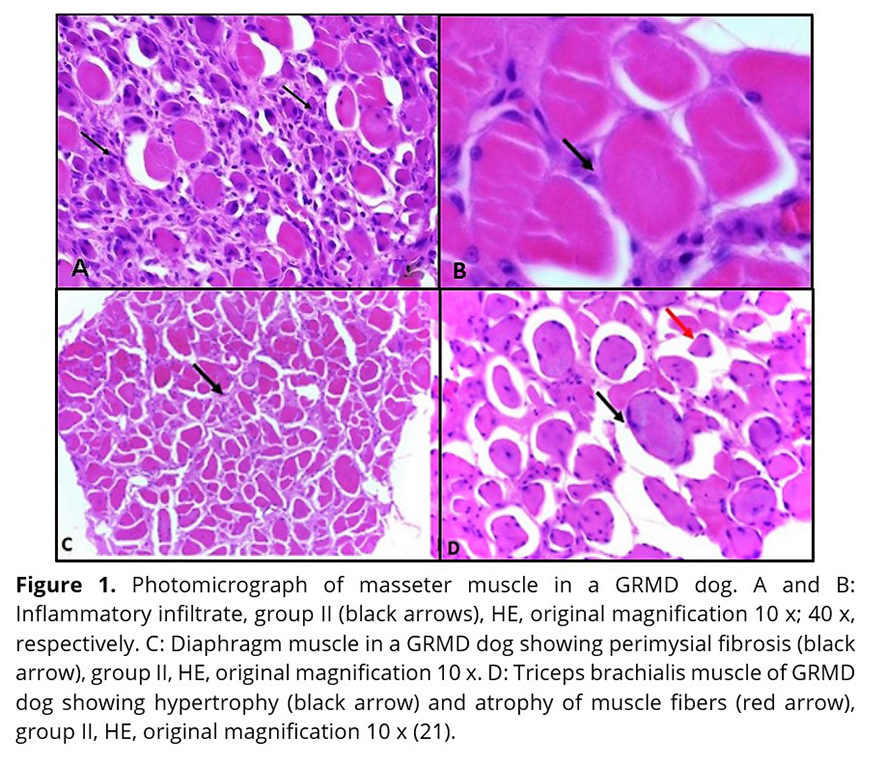
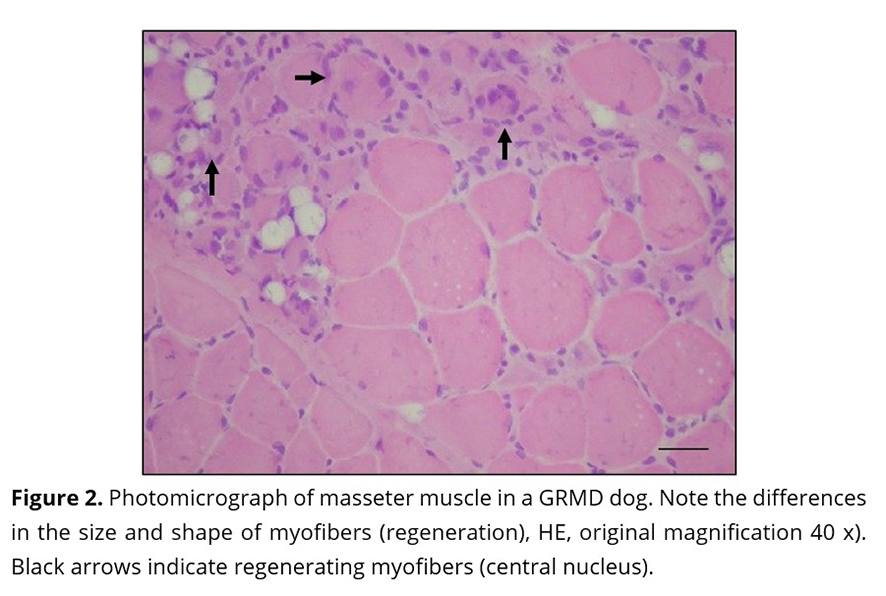
Previous studies have shown inflammatory cell foci (Figure 1 A and B) in all muscles, with more severe lesions in older dogs. The predominant cell group identified was mononuclear (lymphocytes and macrophages)(21). In the control group, inflammatory cell foci were not observed(21). Inflammatory cells were seen in close proximity to the necrotic fibers. Enlargement of the endomysial and perimysial spaces (Figure 1 C) was observed. In all the dystrophic animals, regardless of age, all muscle samples had changes in the diameter of myofibres (hypertrophy and atrophy) with variable degrees of severity (Figure 1D)(21). There was minimal regeneration in the myofibers (Figure 2).
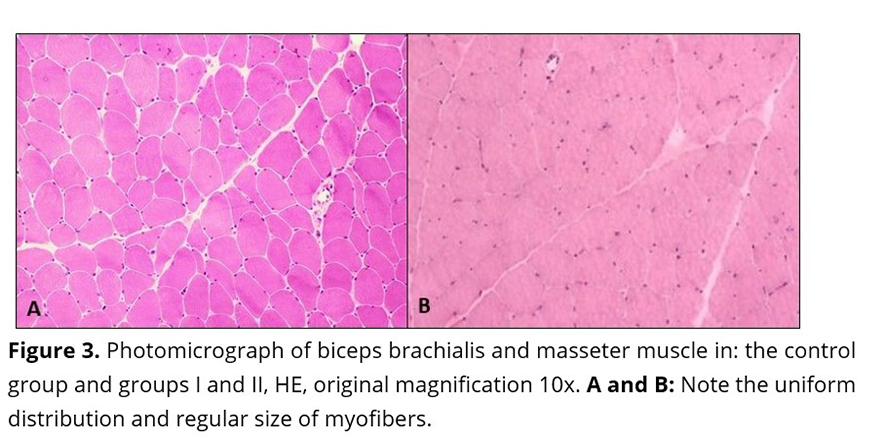
In the control samples, no morphologic or degenerative lesions were described. The samples consisted of cylindrical myofibers and peripheral nuclei. Additionally, the myofibers showed uniform distribution and regular size (Figure 3).
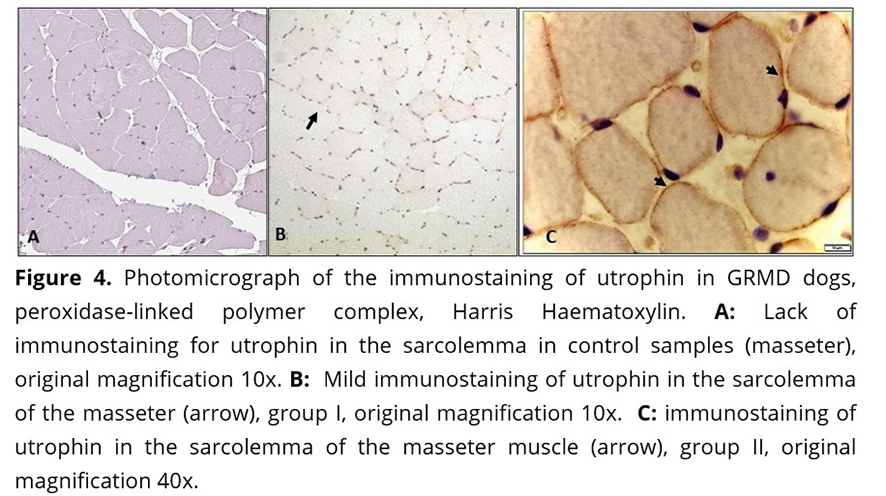
In the dystrophic animals, immunostaining of utrophin was intense (score 3) on the surface of myofibers, blood vessels, and sarcoplasm of some myofibers. In this study, utrophin was expressed in the sarcolemma of dystrophic myofibers of masseter and diaphragm muscles and expression was significantly higher in animals from group II (over one year of age) (Figure 4 B and C) (Table 3). The control group did not show utrophin immunostaining (Figure 4 A).
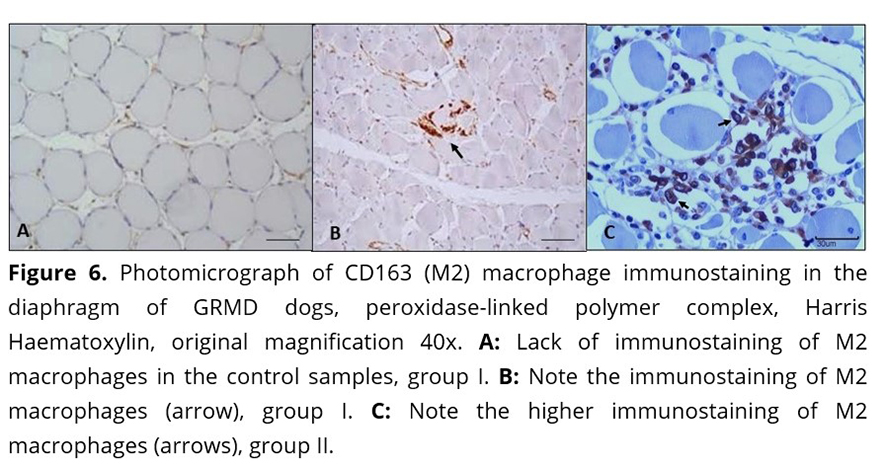
In the muscle samples used as controls, the analysis distribution of macrophages M1 and M2 revealed isolated cells distributed randomly in the endomysium, the perimysium and around vessels (Figure 5A and 6A). These cells were also observed in small groups in the areas of degeneration and necrosis of the dystrophic muscles in the four muscles evaluated.
The distribution CD68 and CD163 immunostaining was different in the four muscles evaluated, showing a direct correlation between the number of macrophages and inflammation grade (Table 1)(21), and Table 4.

There was no significant difference in M1 (CD68) expression in the four muscles evaluated between the two groups of dystrophic animals, indicating that the age of the dystrophic animal had no effect on the number of M1 macrophages. The M1 subtype expressed CD68, which is characteristic of immature lesions. Less immunostaining was seen in necrotic and severely fibrotic areas as shown in Figures 5 B and C.
The immunostaining of M2 macrophages in the two groups of dystrophic animals ranged from moderate to severe in all evaluated muscles. The M2 subtype expressed by CD163 in skeletal muscles of dogs of this study was seen in areas where there was endomysial fibrosis, especially in the diaphragm (Figure 6 B and C) as shown in Table 5. In this muscle, there were twice as many M2 macrophages as M1 macrophages.

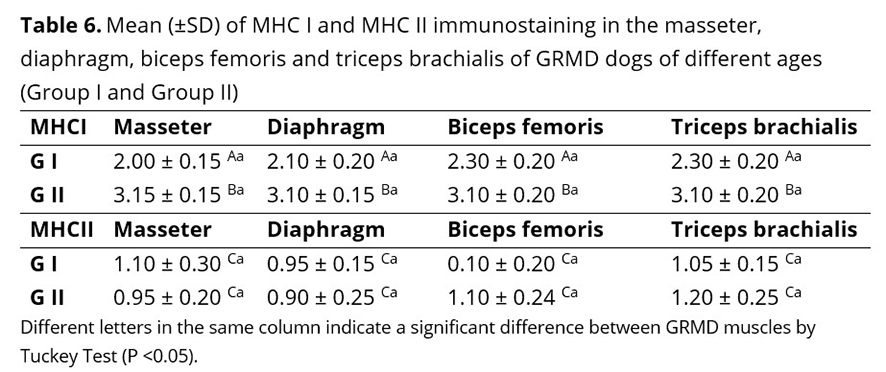
There was expression of MHC I antigen on the surface of dystrophic myofibers and areas of necrosis, regeneration, and inflammation as shown in Figure 7 B and C. MHC I immunostaining differed significantly between the four muscles, although the means of group II immunostaining were higher in all muscles evaluated (Table 6). There was a direct relationship between the intensity of MHCI immunostaining and age, indicating the promoter activity of the MHC I antigen for macrophage performance.
MHC II immunostaining occurred in groups of mononuclear inflammatory cells, especially macrophages, concentrated in areas of degeneration and necrosis, thus confirming their participation in GRMD (Figure 6 E and F). There was no significant difference in MCH II expression in the four muscles evaluated, between the two groups of dystrophic animals, showing that the expression of MHC II did not vary with the age of the dystrophic animal (Table 6).
In control samples, the distribution of MCH I and MHC II revealed isolated immunostained cells distributed randomly throughout endothelium and around vessels.
Discussion
In this study, utrophin expression in the sarcolemma of dystrophic myofiber of masseter and diaphragm and was significantly higher in animals from group II (over one year of age), corroborating findings of other authors who studied the gastrocnemius muscle and the muscle-tendon junction(6). However, our results differed from those who reported that utrophin expression in dystrophin-deficient muscle is characteristic of myofibres that are not regenerating(23). Utrophin immunostaining in DMD patients was weak in mature myofibres of vastus lateralis, gastrocnemius, biceps, quadriceps, lateral rectus and deltoid muscles(7). Due to the intense immunostaining for utrophin observed on the periphery of dystrophic myofibers, it is assumed that it was incorporated into the sarcolemma. These data show the positive regulation of utrophin in the sarcolemma of mature myofibers. The presence of utrophin helps to regulate the function of dystrophin-deficient muscles and provides benefit by the interaction between the actin cytoskeleton and the dystrophin-glycoprotein complex on the sarcolemma. However, it suggests that there is only a small difference in utrophin regulation between different dystrophic muscles in the groups evaluated.
The M1 subtype expressed by CD68 is specific for immature lesions and was found in reduced numbers in necrotic and areas with severe fibrosis. These findings are similar to those described by other authors, who observed weak immunostaining of M1 macrophages compared to M2 subtype in quadriceps muscle from DMD affected patients(24) and soleus muscle in mdx mice(12). This suggests an association of these cells with chronic inflammation in muscle fibers in dystrophic dogs.
There are a number of hypotheses to explain the weak expression of CD68. Firstly, the antibody used for labeling anti-CD68 antigen is not specific to the canine species. Secondly, CD68 is expressed at high levels in peripheral blood and liver, and is not specific for skeletal muscle(25). Thirdly, it is possible that the expression of this antigen in the tissues studied occurs below the sensitivity limit of the immunohistochemical procedure used.
In this study, the CD163 immunostaining of the M2 subtype in canine skeletal muscles was seen in the endomysial areas of the diaphragm and triceps brachialis muscles, especially in the endomysial fibrotic areas. The degree of fibrosis and the intensity of the inflammatory process were proportional in the triceps brachialis and diaphragm muscles(21). GRMD dogs showed a high level of muscle fibrosis when compared to skeletal muscle in healthy dogs. Fibrosis is still poorly understood in dystrophic muscle but appears to be a consequence of the inflammation infiltrate in muscle injury(26). In the diaphragm and triceps brachii muscles, the absolute numbers of M2 macrophages were twice that of M1 macrophages. Studies in the quadriceps muscles of DMD patients showed equivalent expression of M1 and M2(24).
The intense stimulation of Th2 cytokines has been observed in isolated macrophages of mdx mice at one year old, indicating that M2 macrophages expressed receptors for IL-4, IL-10, and IL-13 (most prevalent in this phase of the disease). The development of pathologic fibrosis in skeletal muscle supports data already reported(13, 27, 28, 29). However, the expression of resident macrophages occurs in lower numbers in healthy muscles, reflecting the role of M2C macrophages in the normal physiologic processes in the muscle. Although the number of M2C macrophages can significantly increase in mdx muscle, they remain a small fraction of the total inflammatory cells infiltrating the lesions. In this study, immunostaining of CD163 was similar to that previously described for M2C(13, 29).
Although CD163 expression was higher than that of CD68, it was less than expected for group II where the inflammatory lesions were intense. Therefore, it was concluded that the immunostained cells may not reflect the total number of the cells in the lesion. Due to the intense immunostaining in the regenerating fibers, it was deduced that the main population of macrophages in the lesion was the M2 subtype. These cells act in competition with M1 using arginine and usually reduce the cytotoxic action of M1 cells(6). In this study, there was less competition with M1.
The presence of M2 macrophages in fibrotic areas suggest they play another role in skeletal muscle beyond phagocytosis. Initially, the M2 macrophages accumulated in the epimysium and perimysium and then entered the injured area, after phagocytosis of necrotic tissue was complete. Intense stimulation by Th2 cytokines has been observed in isolated macrophages from mdx mice at one year old, indicating that M2 macrophages express receptors for IL-4, IL-10 and IL-13 (most prevalent in this phase of the disease)(8).
The function of M2 macrophages may be the activation of satellite cells during adaptation of the myofibre and the release of growth factors such as platelet-derived growth factor (PDGF) and fibroblast growth factor (FGF)(30). In other models of muscle cell lesions, the initial activation of satellite cells occurs at the same time as macrophage infiltration into the injured muscle(31). The observation of a smaller number of M1 macrophages (which have inflammatory action and are recruited at sites of acute muscular injury) in areas of necrosis in muscles affected by GRMD confirms the change of their phenotype to M2 macrophages after phagocytosis, showing an anti-inflammatory profile(32, 33). Results of our study supported this finding.
M2 macrophage immunostaining, suggests that there is a moderate increase in this macrophage subtype in muscles of GRMD dogs as they age, indicating the association of this cell type with chronicity of muscle damage in dystrophic dogs.
The number of M2 macrophages (CD163) in affected animals was significantly higher than in control animals, demonstrating the involvement of these cells in GRMD disease, as well as the intensification of MHC I immunostaining with age. This demonstrates the increased expression of surface proteins in the damaged myofibers and the complementary action of M2 macrophages.
The expression of MHC antigen was detected on the surface of dystrophic myofibers and in the necrotic, regeneration and inflammation areas. These data corroborate previous findings that described the expression of this antigen in DMD patients and several muscles (masseter, diaphragm, biceps brachialis, triceps brachialis, semimembranosus, biceps femoris, cranial sartorius, gastrocnemius, and muscle-tendinous junction) from GRMD dogs(6, 34). These features suggest the involvement of macrophages in the pathogenesis of DMD and the activation response to a specific antigen. Although the antigen activation is still unknown, it may result from damage caused by the contraction process in the dystrophic muscle(17).
MHC II immunostaining occurred in mononuclear inflammatory cell groups, especially macrophages, concentrated in areas of degeneration and necrosis confirming their participation in GRMD. These findings were in line with those of previous studies(16, 35).
The macrophage is reported to be the primary cell type involved in dystrophinopathy and its depletion in mdx mice would be responsible for the 80% reduction of muscle necrosis in the early stages of the disease(28). Despite these reports, some authors also found MHC II antigen expression on the surface of dystrophic myofibers(17), a fact not observed in this study. MHC II immunostaining did not vary with the age in this study, in contrast to findings of other authors working with DMD in man(16). However, we cannot exclude the possibility that this antigen was expressed at levels below the sensitivity limit of the immunohistochemical procedure used.
In conclusion, our findings support the hypothesis that M2 macrophages are the main mononuclear inflammatory cells present in lesions in GRMD dogs. The immunostaining of M2 macrophages suggested that in GRMD dogs over one year of age, this macrophage subtype increases in number in the diaphragm. Although this occurred only in the diaphragm muscle sample in this study, we can assume that the same would be true for the other muscles used in this study. This suggests that macrophages are involved in development of chronic muscular lesions of dystrophic dogs.
Conflicts of interest
The authors declare that they are not financially or personally related to any individual or organization that could inappropriately influence or bias the content of this article.
Acknowledgments
The authors would like to thank the Sao Paulo State Research Foundation (FAPESP) for the financial support provided (process number: 11/07906-0).
References