RESUMO
O objetivo deste estudo foi determinar as perdas endógenas e
metabólicas em frangos de corte industrial e caipira em diferentes
idades. No primeiro experimento, utilizaram-se 140 pintos de corte
industrial, distribuídos num delineamento inteiramente casualizado, com
quatro tratamentos e cinco repetições. No segundo experimento, foram
utilizadas 120 aves de sexo misto, da linhagem caipira Embrapa 041,
submetidos a três tratamentos e cinco repetições distribuídas num mesmo
delineamento. Em ambos os experimentos os tratamentos tiveram como
parâmetro as idades médias das aves. Para o experimento 1 as idades
foram 5, 15, 25 e 35 dias de nascimento; para o experimento 2 as idades
médias adotadas foram 15, 45 e 75 dias de nascimento. O período
experimental foi de 72 horas de jejum, sendo as primeiras 24 horas para
limpeza do trato digestivo e as últimas 48 horas de coleta total das
perdas endógenas e metabólicas. Foram determinados os teores de matéria
seca, nitrogênio e energia bruta das perdas endógenas e metabólicas das
aves. Ao se relacionarem as perdas, por ave e peso da ave, diferentes
interpretações foram obtidas para a mesma variável. As equações obtidas
para os frangos industrial e caipira revelaram que essas aves tiveram
comportamentos diferentes na excreção fecal metabólica e urinária
endógena. As perdas diminuíram com o avanço da idade.
PALAVRAS-CHAVES: Aves em jejum, método de coleta total, energia fecal metabólica, energia urinária endógena.
ABSTRACT
AGE EFFECT ON THE ENDOGENOUS AND METABOLIC LOSSES IN BROILER CHICKENS AND FREE RANGE
The objective of this study was to determine the endogenous and
metabolic losses in broiler chickens and free range on different age
rates. On the experiment 1, 140 broiler chickens distributed in
an entirely randomized design with four treatments and five
replications have been used. On the second experiment, 120 birds of
mixed sex from the Embrapa 041 free range line distributed in tree
treatments and five replications distributed in a same design. On both
experiments the treatments consisted in the average ages of the birds.
To the first experiment the ages were: 5; 15; 25 and 35 days of age and
the second experiment the average ages adopted were 15, 45 and 75 days
of age. The experimental period was of 72 hours of fasting, being
the first 24 hours for digestive tract cleaning and the last 48 hours
for total collection of the endogenous and metabolic losses. The
contents of dry matter, nitrogen and brute energy from birds`
endogenous and metabolic losses have been determined. When relating
losses per bird and bird weight, different interpretations were
obtained for the same variable. The equations obtained to broiler
chicken and free range reveled that those birds had different behaviors
during metabolic fecal and endogenous urinary. The losses diminished
according to age advance.
KEYWORDS: Birds on fasting, total collection method, metabolic fecal energy, endogenous urinary energy.
INTRODUÇÃO
Há mais de três décadas pesquisadores estudam as perdas endógenas e
metabólicas e seus respectivos efeitos sobre a determinação dos valores
energéticos dos alimentos para aves. Nesse período, aprimoraram-se o
conceito, e a padronização das metodologias, a fim de aumentar a
precisão na determinação real de energia aproveitada pelos animais.
Conceitualmente, de forma detalhada, nas fezes das aves encontram-se os
resíduos não digeridos e a fração metabólica formada por bílis,
secreções digestivas e células procedentes da mucosa intestinal e nas
aves não cecectomizadas à ação da microbiota. Na fração urinária
predominam os resíduos alimentares absorvidos e eliminados sem ser
catabolizados, adicionados da fração endógena, que consiste no produto
do catabolismo dos tecidos (SIBBALD & PRICE, 1978; SIBBALD, 1981a;
SILVA
et al., 2006).
O sistema de energia metabolizável aparente (EMA) supõe que toda
energia excretada é proveniente do alimento, o que não é verdade.
Motivado por essa premissa, SIBBALD (1976) propôs a correção das perdas
endógenas e metabólicas nos valores de EMA, sugerindo que o sistema de
energia metabolizável verdadeira (EMV) representaria melhor a energia
do alimento.
O referido autor afirmou que as frações endógenas e metabólicas presentes na excreta das aves em regime a
d libitum
são semelhantes às frações coletadas em aves submetidas ao jejum. Essa
afirmação também não é verdadeira, e tem motivado discussões no campo
teórico entre os pesquisadores.
Assim, na literatura existem relatos de que o sistema de EMV expressa
com mais segurança o conteúdo energético dos alimentos para aves,
quando comparado ao sistema de EMA (SIBBALD, 1976; DALE & FULLER,
1982; BORGES
et al., 2003). Contudo, essa premissa não tem sido confirmada em alguns trabalhos de desempenho animal, como o de FREITAS
et al. (2006), que verificaram maiores consumo de ração, ganho de peso e melhor conversão alimentar com o sistema de EMA.
Hoje, sabe-se que dentre os fatores que afetam a intensidade das perdas
endógenas e metabólicas em aves estão: a metodologia aplicada, o regime
alimentar, a genética, a categoria (BORGES
et al., 2003) e, para frangos industriais, como descoberto recentemente, especificamente a idade (SILVA
et al.,
2006). Dessa maneira, o objetivo deste trabalho de pesquisa foi
determinar o efeito da idade sobre as perdas endógenas e metabólicas em
frangos de corte industrial e caipira.
MATERIAL E MÉTODOS
Realizaram-se dois experimentos: um com frango de corte industrial e
outro com frango de corte caipira. O primeiro experimento foi feito no
Departamento de Zootecnia da Universidade Federal Rural de Pernambuco
(UFRPE), com 140 pintos de corte machos da linhagem Cobb, de um dia de
idade. As aves foram distribuídas em um delineamento experimental
inteiramente casualizado, com quatro tratamentos e cinco repetições.
Os tratamentos consistiram em quatro idades médias de frango de corte
(5, 15, 25 e 35 dias de idade), sendo 10, 8, 6 e 4 o número de aves por
parcela experimental nas respectivas idades. Os pesos médios das aves
no início do período experimental foram 83,28g (T1); 434,10g (T2);
1.004,80g (T3) e 2.310,60g (T4). As aves foram selecionadas pelo peso,
buscando a máxima uniformidade, e alojadas em gaiolas de metabolismo
dotadas de sistema de aquecimento na primeira semana de vida e de
bandejas coletoras sob o piso.
Inicialmente, transferiram-se as aves para as baterias (1,00 x 0,50 x
0,50 m) nos 1º, 10º, 20º e 30º dias de idade, obedecendo-se a um
período de 4 dias de adaptação às instalações. As aves foram mantidas
em jejum a partir dos 4º, 14º, 24º e 34º dias por um período de 24
horas para esvaziar o trato digestório e, em seguida, por mais 48
horas, para coleta do material, resultando em idades médias
experimentais ao 5º dia de idade (T1), 15º dia de idade (T2), 25º dia
de idade (T3) e 35º dia de idade (T4), como já relatado.
As aves utilizadas foram descartadas ao final de cada experimento.
Assim, de um total de 240, para cada idade selecionaram-se novas aves
que foram alojadas no primeiro dia de vida em boxes dotados de lâmpadas
incandescentes de 100 watts para aquecimento nas duas primeiras semanas
de vida. O material utilizado como cama foi maravalha. As aves foram
arraçoadas com uma dieta à base de milho e farelo de soja, formulada
para atender às exigências nutricionais nas diferentes idades, segundo
ROSTAGNO
et al. (2005).
Durante o experimento, após o jejum de 24 horas em cada idade, fez-se
duas vezes ao dia a coleta dos resíduos excretados (material endógeno e
metabólico) pelas aves por um período de 48 horas. Em seguida, o
material coletado de cada parcela experimental foi acondicionado e
congelado em freezer a -20ºC.
Ao término do experimento, o material foi descongelado, homogeneizado,
amostrado, pesado e pré-seco em estufa de ventilação forçada regulada
para 55ºC por 72 horas. No fim da pré-secagem, as excretas foram moídas
em moinho de facas, dotado de peneira de 1 mm de abertura de crivo,
para serem feitas as análises de matéria seca, nitrogênio e energia
bruta, seguindo as metodologias descritas por SILVA & QUEIROZ
(2002).
Após as análises, calcularam-se a matéria seca fecal metabólica e
urinária endógena (MSfm + MSue), o nitrogênio fecal metabólico e
urinário endógeno (Nfm + Nue) e a energia fecal metabólica e urinária
endógena (Efm+Eue), além das perdas endógenas e metabólicas de matéria
seca e nitrogênio.
O segundo experimento foi conduzido na Estação Experimental de Pequenos
Animais de Carpina (EEPAC), pertencente à UFRPE. Utilizaram-se 120 aves
de sexo misto da linhagem caipira Embrapa 041. As aves foram
distribuídas num delineamento experimental inteiramente casualizado,
com três tratamentos e cinco repetições. Os tratamentos tiveram como
parâmetros as idades médias dos frangos caipiras: 15, 45 e 75 dias de
idade, utilizando dez, oito e seis aves por parcela experimental nas
respectivas idades, sendo metade de cada sexo.
As aves foram transferidas para as baterias (1,00 x 0,50 x 0,50 m) aos
10º, 40º e 70º dias de idade e obedeceu-se a um período de 4 dias de
adaptação às instalações. As aves foram mantidas em jejum aos 14º, 44º
e 74º dias por 24 horas, para limpar todo trato digestório; em seguida
houve jejum por mais 48 horas, para coleta do material, resultando em
idades médias experimentais aos 14 dias de idade (T1), 45 dias de idade
(T2) e 75 dias de idade (T3).
No período pré-experimental, as aves foram criadas em galpão
convencional, com fornecimento de ração e água à vontade. A ração
ofertada era à base de milho e farelo de soja, formulada de acordo com
as exigências nutricionais para as diferentes idades, como preconizado
pelo manual da linhagem (
Tabela 1).
A seleção dos animais foi baseada no peso médio das aves, buscando a
uniformidade da parcela. Os pesos médios foram 0,273g, 0,921g e 2,047g
para as idades de 15, 45 e 75 dias, respectivamente. Em ambos os
experimentos forneceu-se água à vontade durante todo o período. A
coleta, manipulação, análise do material excretado e os cálculos dos
parâmetros interessados foram os mesmos citados no primeiro
experimento.
As variáveis dependentes analisadas foram: perdas endógenas e
metabólicas de matéria seca, nitrogênio e energia bruta, nas diferentes
idades, expressas em diferentes sistemas de unidade. Os resultados
obtidos foram submetidos à análise de regressão, utilizando-se o
programa computacional SISVAR versão 4.6 (FERREIRA, 2003).
RESULTADOS E DISCUSSÃO
A composição química das perdas endógenas e metabólicas de frangos de corte industrial e caipira está apresentada na
Tabela 2.
Os resultados encontrados revelaram que a maior concentração de matéria
seca (MS) foi verificada a partir dos 25 dias de idade, enquanto a
menor concentração encontrava-se nas aves com idade média de 5 dias.
Diferentemente da MS, a energia bruta (EB) das perdas endógenas e
metabólicas não teve alterações nas diferentes idades; a análise de
nitrogênio (N) apresentou o menor teor aos 25 dias de idade, não
existindo diferenças entre as demais idades.
As perdas endógenas e metabólicas dos frangos de corte caipira tiveram
as maiores concentrações de matéria seca e energia bruta aos 45 dias de
idade. Nessa idade a concentração de nitrogênio apresentou o menor
teor, não existindo diferenças entre as idades equidistantes (15 e 75
dias). Fatores relacionados a jejum, idade, composição corporal,
genótipo e estado fisiológico da ave são relatados (SIBBALD, 1981b;
SILVA
et al., 2006) como
condicionantes para a composição química das perdas endógenas e
metabólicas. Assim, a composição química deve ser relacionada a outras
variáveis, para que faça a contextualização adequada e para que haja
interpretações conclusivas e coerentes sobre os acontecimentos pelos
quais a ave passou em estado de jejum e em jejum prolongado.
Ao se relacionar a matéria seca fecal metabólica e urinária endógena
com o volume excretado por parcela, por ave e por peso da ave, nota-se
que as diferenças entre as idades existem (p≤0,01); contudo, conforme o
sistema em que ela está sendo expressa, os resultados revelam
comportamentos opostos (
Tabela 3).
Ao se fazer a relação com o peso corporal da ave, observou-se um
resultado oposto. Assim, as aves com a idade média de 5 dias excretaram
uma quantidade equivalente a 8% do seu peso. As diferenças se anularam
a partir do 15º dia de vida e ao 35o dia a perda correspondeu a pouco
mais de um décimo do peso corporal da ave.
O ajuste do metabolismo às diferentes condições fisiológicas é obtido
graças aos processos que compõem a regulação metabólica. Assim, 12
horas após a refeição inicia-se o estado pós-absortivo; passado esse
momento, a degradação de glicogênio, a proteólise muscular e a lipólise
se sucedem, sendo responsáveis por manter o aporte energético no
organismo (MALHEIROS, 2008).
No momento de jejum, o glucagon é favorecido na relação
insulina-glucagon, e a glicogenólise é intensificada. Já no jejum
prolongado, o índice do glicêmico é mantido pela gliconeogênese,
relacionada à perda significativa de massa muscular e de tecido adiposo
que acompanham o jejum (NELSON & COX, 2000).
De forma prática, a relação entre o metabolismo basal e o peso
metabólico possibilita justificar e interpretar mais coerentemente os
eventos fisiológicos ocorridos na ave, sobretudo quando se compara a
taxa metabólica de animais com pesos e composição corporais diferentes,
conforme BLAXTER (1989).
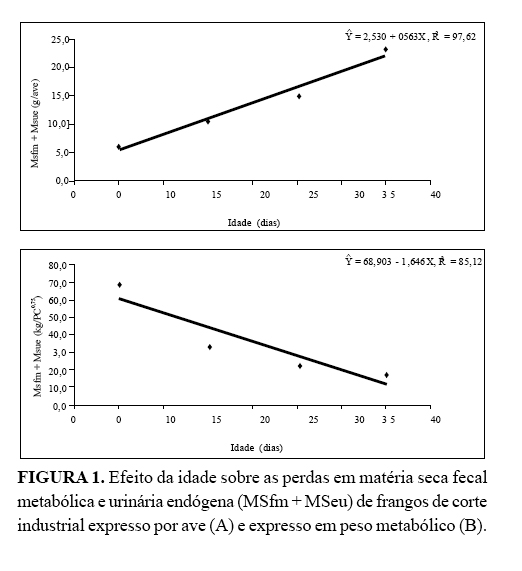
Assim, a grama de matéria seca fecal metabólica e urinária endógena
(MSfm + MSeu) excretada por ave aumentou com o avançar da idade de
forma linear Ŷ = 2,530 + 0,563X, R2 = 97,62 (
Figura 1A).
Contrariamente quando se relacionou a excreção de MSfm + MSeu ao peso
metabólico da ave, a excreção diminuiu cerca de 1,646 g/dia, conforme a
equação Ŷ = 68,903 - 1,646X, R2 = 85,12 (
Figura 1B).
Nessa fase são marcantes as diferenças nos constituintes químicos corporais da ave (SILVA
et al.,
2006), e esses fatores são responsáveis pelas diferenças apresentadas,
uma vez que a composição corporal se relaciona intimamente com a
expressão e a intensidade das perdas endógenas e metabólicas, segundo
os autores supracitados.
A
Tabela 4
mostra diferenças marcantes na fase inicial das aves de crescimento
lento, em que a excreção MSfm + MSeu por ave foi aproximadamente 2,4
vezes menor que a excreção nas fases subsequentes, crescimento e final.
Na fase inicial, comparativamente, 4,25 g de matéria seca faz diferir o
quantitativo expelido pelas aves de corte industrial e pelas aves de
corte caipira. Em relação percentual, as aves de corte industrial
perdem no mesmo período de coleta cerca de 70% a mais que as aves
caipiras.
Assim como as aves de corte industrial, as caipiras também apresentaram
comportamento inverso quando se relacionou a variável grama de matéria
seca com o peso corporal e com o peso metabólico. Diferentemente do
sistema anterior (gMS/ave), os sistemas gMS/kgPC, % de PC e gMS/kgPC
0,75
exibiram diferenças (p<0,01) entres as idades experimentais. Para o
sistema gMS/kgPC, entre o 15º e o 45º dia de idade a redução observada
foi de cerca de 27%. Esse percentual aumentou entre as idades
subsequentes, (45º e 75º dias de idade), chegando próximo a 58% de
redução, até atingir 1,97 gMS/kgPC na última idade avaliada.
O efeito da idade de frangos caipiras sobre as perdas endógenas de matéria seca metabólica e urinária pode ser visto nas
Figuras 2A e
2B
de forma mais nítida, sobretudo quando se relaciona MSfm + MSeu em
quilo de peso metabólico. Dessa forma, a excreção por ave diminuiu em
0,073 g por dia até a última idade experimental (
Figura 2B), em detrimento da excreção no sistema grama ave (
Figura 2A),
que apresentou aumento expressivo até o 45o dia de idade. Entre essa
idade e a subsequente as diferenças não foram percebidas pelo teste de
média, e a análise de regressão mostrou um modelo (Ŷ = 1,675 + 0,037X,
R2= 65,79) com grau de dispersão considerável, comparado ao modelo
anunciado anteriormente com frangos de corte industrial.
Esse aumento significativo até o 45º dia pode ser relacionado à idade
fisiológica da ave, ou seja, nessa idade a taxa de crescimento começa a
diminuir significativamente (ponto de inflexão da reta) e leva a uma
diminuição na taxa metabólica da ave. Ensaios de crescimento com aves
dessa categoria e grupo genético foram realizados por SANTOS
et al.
(2005). Esses autores verificaram, por meio da aplicação da equação de
Gompertz, que no ponto de mudança do padrão de aceleração na curva – ou
seja, o ponto de inflexão, em que a ave atinge a maior taxa de
crescimento, por volta do 46º dia de vida – a atividade metabólica é
intensa.
Esse estado fisiológico pode ter influenciado o aumento expressivo
percebido até os 45 dias de idade. Após esse período, ou seja, na fase
subsequente, não se observaram alterações marcantes na excreção
metabólica e endógena de matéria seca por ave analisada no presente
experimento.
Na
Tabela 5
estão apresentados os dados referentes ao nitrogênio metabólico e
endógeno (Nfm + Nue) quantificado em frangos de corte industrial em
diferentes idades. Os dados obtidos sugerem que as maiores diferenças
na excreção do Nfm + Nue ocorrem nas idades equidistantes 5 e 35 dias,
independentemente da forma como está sendo expressa. Desse modo, a ave
aumenta sua excreção em 1,112 g por dia.
Essa informação, relacionada ao peso corporal, indica que
proporcionalmente a maior perda acontece na fase pré-inicial e, após
esse período, há redução de cerca de 0,42 g por dia. Assim, o
nitrogênio endógeno e metabólico quantificado na excreta de aves com 15
dias de idade reduz cerca de 66% quando comparado com a menor idade. Em
seguida, de 15 a 25 dias de idade, essa redução diminui para 54% e
tende a estabilizar entre 25 e 35 dias de idade, chegando a apenas
12,5%, de redução.
A perda de nitrogênio no trato digestório tem motivado estudos há
bastante tempo. A secreção endógena de nitrogênio do trato digestivo de
ratos foi investigada pelos pesquisadores TWOMBLY & MEYER (1961),
utilizando ensaios de curta duração. Nesses ensaios, os referidos
autores observaram que à medida que aumentava a percentagem de proteína
da dieta aumentava o teor de nitrogênio em 0,0459 g, conforme a equação
Ŷ = 1,31 + 0,0459X. Para o nível máximo de proteína na dieta ao final
das 12 horas de alimentação, esses pesquisadores estimaram que cerca de
2,228 g de nitrogênio foram secretados pelo epitélio secretor do trato
digestório. Por extrapolação ao consumo zero de proteína, para dieta
isenta de proteína, TWOMBLY & MEYER (1961) verificaram algo próximo
de 1,31 g de nitrogênio nas fezes dos animais.
Dados coletados por CHUNG & BAKER (1992), utilizando galos adultos,
revelaram que a perda de nitrogênio fecal metabólico e urinário
endógeno (Nfm + Nue) foi próxima de 0,944 g por ave. Dados recentes
publicados por SILVA
et al.
(2006) referentes a aves mantidas em jejum demonstraram que um pinto de
corte industrial ao 7o dia de vida perde em média 0,61 g de Nfm + Nue;
o mesmo frango ao 37º dia aumenta essa excreção cerca de 3,5 vezes.
Esse aumento, nessa mesma proporção, ocorreu nos valores obtidos no
presente trabalho.
No entanto, os números absolutos registrados neste estudo, nas
diferentes idades, foram superiores aos valores reportados por SILVA
et al. (2006). Isso se pauta nos aspectos metodológicos, no que se refere à idade e ao peso da ave submetida ao jejum experimental.
Na
Tabela 6
estão apresentados os valores das perdas de nitrogênio pelas aves
caipiras. Pode ser visto que a excreção de nitrogênio aumentou com o
avanço da idade numa proporção de 0,007 g por dia, sendo o maior
aumento entre 15 e 45 dias de idade. A diferença na excreção entre 15 e
45 dias foi de 1,97 vezes – aumento esperado, uma vez que o comprimento
do intestino cresce sensivelmente nesse intervalo, conforme foi
verificado por FIGUEIREDO
et al. (2002).
SANTOS
et al. (2005)
verificaram que a relação dos intestinos com o peso do animal em jejum
corresponde a 3,35% em frangos de corte caipiras e a 3,8% nos frangos
de corte industriais. Em números relativos essa diferença é estreita,
mas em valores absoluto, o frango de corte industrial dispõe de uma
maior superfície intestinal para ocorrências de descamações celulares.
Essa característica acentuou as diferenças nos resultados obtidos, ao
passo que o valor médio verificado para os frangos de corte foi cerca
de 4,6 vezes maior que o obtido com as aves caipiras.
A fim de atender às necessidades corporais com o avançar da idade, para
desenvolver e aprimorar a capacidade digestiva-absortiva, o crescimento
dos intestinos obriga a ocorrência de modificações no epitélio,
especialmente no jejuno e íleo, no que se refere ao número e ao volume
dos enterócitos (MORAN JUNIOR, 1985; UNI
et al.,
1995). Esses fenômenos resultam em descamações diárias que, somadas à
flora do intestino grosso e à mobilização de reservas corporais para
atender à exigência de mantença no período de jejum, podem ter exercido
efeito direto sobre o quantitativo de nitrogênio presente nas perdas
endógenas e metabólicas, mais intensas frango de corte industrial.
No estado de jejum prolongado, o nível de aminoácidos circulantes na
corrente sanguínea é alto. Esse, por sua vez, são direcionados às
células do tecido hepático, onde sofrem oxidação completa ou mesmo
podem ser convertidos em glicose, bem como em corpos cetônicos
(BROSNAN, 2000; MALHEIROS, 2008). Nesse momento, o aminoácido alanina
desempenha papel crucial como importante substrato da gliconeogênese
(STRAYER, 1995; NELSON & COX, 2000). Segundo esses estudiosos, as
proteínas do tecido muscular são utilizadas para essa finalidade, mas
num período curto, pois tende a ser preservada, uma vez que sua reserva
é limitada.
Assim, a mobilização de reservas é a principal fonte para síntese de
glicose hepática durante períodos de jejum prolongado; os precursores
prioritários são aminoácidos, advindos principalmente do músculo
esquelético. Em outro momento, o glicerol proveniente da mobilização
das reservas de gordura, triglicerídeos do tecido adiposo e o lactato
oriundo das hemáceas constituem os artifícios utilizados pelo
metabolismo do animal durante a coleta de dados utilizando essa
metodologia, com aves em jejum.
Dessa forma, aves em regime ad libitum utilizam artifícios metabólicos
diferentes dos anunciados anteriormente. Assim, as perdas endógenas e
metabólicas tendem a ser diferentes, uma vez que o nitrogênio presente
na urina (em aves alimentadas) não vem da mobilização de reservas, e
sim de catabolismo dos aminoácidos provindos da dieta.
Informações dessa natureza, segundo WARPECHOWSKI
et al.
(2006), têm sido motivo de discussão em relação ao sistema de energia
metabolizável verdadeira, uma vez que no cálculo do balanço de
nitrogênio verdadeiro se computa a perda endógena e metabólica desse
componente. Dessa maneira, o balanço é influenciado diretamente pelo
método utilizado na coleta do dado e, consequentemente, nos valores de
energia metabolizável.
Nas
Tabelas 7 e
8
estão apresentadas as perdas de energia seca fecal metabólica e
urinária endógena (Efm + Eue) em frangos de corte industrial e caipira
em função da idade. A excreção de energia fecal metabólica e urinária
endógena por parcela mostrou-se diferente apenas na fase pré-inicial,
não havendo mais alteração nas fases subsequentes, conforme a análise
estatística. No entanto, é prudente salientar que alto coeficiente de
variação fez que a diferença mínima significativa fosse superior a 43
kcal nas demais idades.
Os modelos de equações gerados para a linhagem industrial foram
lineares, enquanto as aves caipiras se adequaram aos modelos
quadráticos. A Efm + Eue verificada para cada frango de corte caipira
assumiu o valor máximo de 15,7 kcal aos 57 dias de idade, conforme o
modelo Ŷ = -2,79 + 0,65X - 0,0057X2. A excreção máxima das aves
caipiras foi inferior ao menor valor obtido para os frangos de corte
industrial, que segundo a análise de regressão aumenta conforme a idade
em 1,86 kcal por dia.
A energia metabolizável verdadeira (EMV) é obtida pela diferença entre
a energia consumida (Econ) e a energia excretada (Eexc), corrigida para
energia fecal metabólica e urinária endógena (Efm + Eeu). Assim, a
fórmula é: EMV = Econ – Eexc – MSfm + MSeu. Admitindo-se consumo de
ração e excreção constantes, variando-se apenas o valor da Efm + Eeu
aos 5 dias (208 kcal) e a média (308 kcal) entre os 15 e os 35 dias
idade, no cálculo da EMV, pode se especular que a adoção do valor médio
de 308 kcal/parcela pode superestimar metabolizabilidade de uma ração
para pintos com 5 dias de idade em mais de 8%, considerando como fonte
de variação unicamente a Efm + Eeu para pintos de corte industrial.
Isso se deve ao fato de que, em baixos níveis de consumo, as perdas são
proporcionalmente maiores e resultam em diminuição dos valores
estimados para energia metabolizável aparente (EMA) e aparente
corrigida (EMAn), segundo LIMA
et al. (1989).
SILVA
et al. (2006) ressaltam
que efeitos derivados a partir do baixo consumo por aves experimentais
são mais pronunciados quando se utiliza a metodologia de alimentação
forçada preconizada por SIBBALD (1976). Resultados de experimentos
conduzidos por LIMA
et al.
(1989) apontaram para valores significativamente diferentes quando
comparados aos valores de EMV vs EMA e EMVn vs EMAn, evidenciando o
efeito das perdas endógenas e metabólicas em baixos níveis de ingestão.
Em níveis normais de consumo, as perdas endógenas e metabólicas são
pequenas em relação à excreção de energia proveniente do alimento e
tendem a demonstrar pouca influência nos valores de EMA e EMAn (LIMA
et al., 1989).
Embora alguns trabalhos tenham visto que matematicamente não há
diferença nos valores de energia metabolizável utilizando a metodologia
tradicional de coleta total com aves em regime ad libitum, a natureza
dos compostos excretados e o perfil metabólico das aves em jejum são
diferentes dos encontrados nas aves sob alimentação. Assim, a correção
feita nos valores de EMV com informações das perdas endógenas e
metabólicas de aves mantidas em jejum não representa o real valor da
energia do alimento. Além desses aspectos, elas ainda diferem com a
idade e o peso metabólico, seja frango industrial ou caipira.
CONCLUSÃO
As perdas endógenas e metabólicas de frangos de corte industrial e
caipira diminuíram com o avanço da idade, sendo mais acentuadas para
frangos de corte industrial.
REFERÊNCIAS
BLAXTER, K. Energy metabolism in animals and man. Cambridge:
Cambridge University Press, 1989. 336 p.
BORGES, F. M. O.; ROSTAGNO, H. S.;
SAAD, C. E. P.; RODRIGUEZ, N. M.; TEIXEIRA, E. A.; LARA, L. B.; MENDES, W. S.,
ARAÚJO, V. L. Comparação de métodos de avaliação dos valores energéticos do
grão de trigo e seus subprodutos para frangos de corte. Arquivo Brasileiro
Medicina Veterinária e Zootecnia, v. 55, n. 6, p. 710-721, 2003.
BROSNAN, .J. T. Glutamate, at the interface between amino acid and carbohydrate
metabolism. Journal of Nutrition, v. 130, n. 4, p. 988S-990S, 2000.
CHUNG, T. K.; BAKER, D. H. Apparent and true
amino acid digestibility of a crystalline amino acid mixture and of casein:
comparison of values obtained with ileal-cannlated pigs and cecectomized
cockerels. Journal Animal Science, v. 70, n. 12, p. 3.781-3.790, 1992.
DALE, N. M.; FULLER, H. L. Applicability of the metabolizable energy
system in practical feed formulation. Poultry Science, v. 61, n.
2, p. 351-356, 1982.
FERREIRA, D. F. Programa SISVAR. Sistema
de análise de variância. Versão 4.6. Lavras: Universidade Federal de
Lavras, 2003.
FIGUEIREDO, E. P. A.; ROSA, O. S.;
OMM, E. R.; BOFF, J. A.; BASSI, L. Peso e tamanho dos órgãos de diferentes
linhagens de frangos de corte. Revista Brasileira de Ciência Avícola, v.
4, supl. 4, p. 95, 2002.
FREITAS, E. R.; SAKOMURA, N. K.;
EZEQUIEL, J. M. B.; NEME, R.; MENDONÇA, M. O. Energia metabolizável de
alimentos na formulação de ração para frangos de corte. Pesquisa
Agropecuária Brasileira, v. 41, n. 1, p. 107-115, 2006.
LIMA, L. I.; SILVA, J. D. da.;
ROSTAGNO, S. H. TARFURI, M. L. Composição química e valores energéticos de
alguns alimentos determinados com pintos e galos, utilizando duas metodologias.
Revista Brasileira de Zootecnia, v. 18, n. 6, p. 546-556, 1989.
MALHEIROS, S. V. P. Integração
metabólica nos períodos pós-prandial e de jejum. Revista Brasileira de
Ensino de Bioquímica e Biologia Molecular. Disponível em:
<http://www.bdc.ib.unicamp.br/rbebbm/>. Acesso em: 22 jul. 2008.
MORAN JR. E. T. Digestion and absorption of carbohydrates in fowl and
events through perinatal development. Journal of Nutrition, v. 115, n.
5, p. 665-674, 1985.
NELSON, D. L.; COX, M. M.; Lehninger, principles of biochemistry.
3. ed. Worths Publishers: New York, 2000. p. 949.
ROSTAGNO, H. S.; ALBINO, L. F. T.;
DONZELE, J. L.; GOMES, P. C.; OLIVEIRA, R. F. M.; LOPES, D. C.; FERREIRA, A.
S.; BARRETO, S. L. T. Tabelas brasileiras para aves e suínos (Composição de
alimentos e exigências nutricionais). 2. ed. Viçosa: Edutira UFV, 2005. 186
p.
SANTOS A. L. dos.; SAKOMURA, N. K.;
FREITAS, E. R.; FORTES, C. M. L. S.; CARRILHO, E. N. V. M.; FERNANDES, J. B. K.
Estudo do crescimento, desempenho, rendimento de carcaça e qualidade de carne
de três linhagens de frango de corte. Revista Brasileira de Zootecnia,
v. 34, n. 5, p. 1.589-1.598, 2005.
SIBBALD, I. R. Metabolic plus endogenous energy
and nitrogen loses of adult cockerels the correction used in the bioassay for
true metabolizable energy. Poultry Science, v. 60, n. 4, p. 805-811,
1981a.
SIBBALD, I. R. Metabolic plus endogenous energy excretion by fowl. Poultry
Science, v. 60, n. 12, p. 2.672-2.677, 1981b.
SIBBALD, I. R. The true metabolizable energy values of several
feedstuffs measured with roosters, laying hens, turkeys and broiler hens. Poultry
Science, v. 55, n. 4, p. 1.459-1.463, 1976.
SIBBALD, I. R.; PRICE, K. The metabolic and endogenous energy losses of
adult roosters. Poultry Science, v. 57, n. 2, p. 556-557, 1978.
SILVA, D. J.; QUEIROZ, A. C. Análise
de alimentos (métodos químicos e biológicos). 3. ed. Viçosa: Editora UFV,
2002. 235 p.
SILVA, E. P.; RABELLO, C. B. V.;
LIRA, R. C.; FARIAS FILHO, R. V.; ALBUQUERQUE, C. S.; SILVA, da. T. Estimativas
das perdas endógenas e metabólicas em frangos de corte. Revista Brasileira
de Ciências Agrárias, v. 1, n. único, p. 115-121, 2006.
STRAYER,
L. Biochemistry. 4. ed. New York: W.H. Freeman, 1995.
TWOMBLY, J.; MEYER, J. H. Endogenous nitrogen
secretions into the digestive tract. Journal of Nutrition, v. 74, n. 4,
p. 453-460, 1961.
UNI, Z.; NOY, Y.; SKLAN, D. Post-hatch changes in morphology and
function of the small intestine in heavy and light strain chicks. Poultry
Science, v. 74, n. 10, p. 1.622-1.629, 1995.
WARPECHOWSKI,
M. B.; KESSLER, A. M. POPHAL, S.; EBERT, A.; RIBEIRO, A. M. L. Digestibilidade
ideal verdadeira da proteína em frangos de corte sob dietas com diferentes
níveis de proteína bruta. Acta Scientiarum Animal Science, v. 28, n. 3,
p. 281-287, 2006.
Protocolado
em: 21 nov. 2008. Aceito em: 30 set.
2010.
{kind=link}