
Introdução
A redução no custo da produção animal deve ser direcionada para a utilização racional de todos os recursos alimentares disponíveis, assim, o setor produtivo vem buscando alternativas de menor custo para evitar perdas na produtividade dos rebanhos(1). O uso de espécies nativas como fonte de volumoso em períodos de déficit forrageiro torna-se uma prática comum na região Nordeste, onde o período seco compromete os índices produtivos nesses sistemas. No entanto, o uso dessas espécies é limitado, muitas vezes, por seu baixo valor nutritivo, o que leva ao menor aproveitamento dos nutrientes pelos microrganismos ruminais(2).
O uso de tratamentos químicos como forma de melhorar a qualidade de forrageiras nativas é uma das formas de otimizar sua utilização em sistemas de alimentação de ruminantes. Dentre os tratamentos utilizados, destaca-se a amonização com ureia, que melhora o aproveitamento da fração fibrosa por hidrolisar as ligações covalentes tipo éster entre a lignina e a parede celular, além de fornecer NNP na forma de amônia(3).
Dentre as forrageiras nativas com potencial de uso em dietas para ruminantes no Meio-Norte do Brasil, destacam-se as plantas jovens de babaçu, conhecidas popularmente como pindobas e que ocorrem em sucessão secundária em áreas de pastagens nesta sub-região(4). O uso da folhagem de babaçu jovens como suporte forrageiro, além de melhorar a oferta de volumoso, reduz o desmatamento de áreas babaçuais para implantação de pastagens cultivadas, prática comum decorrente da desinformação do seu valor nutritivo. Além disso, o corte das folhagens de babaçu jovens está de acordo com a lei 231B/2017 que regula o corte das palmeiras, impedindo seu desbaste total em áreas estabelecidas por plantas adultas.
A eficiência da amonização na melhoria da qualidade de volumosos foi constatada por Oliveira et al.(5) ao avaliarem a cana de açúcar amonizada com 6% de ureia, com reduções de 13,87% na fração FDN e por Moreira Filho et al.(6) com redução de 6,4% nessa fração para o feno do restolho de milho. Assim, objetivou-se com esta pesquisa avaliar a composição química e a degradação in situ do feno de pindoba de babaçu submetido à amonização com ureia em duas alturas.
Material e métodos
O experimento foi realizado no setor de caprinocultura da Universidade Federal do Piauí, no município de Teresina, e as folhas de pindobas de babaçu foram colhidas em área de ocorrência natural da espécie no setor, sendo plantas em sucessão secundária, após o corte para implantação de pastagem cultivada. O trabalho foi submetido à avaliação da Comissão de Ética no Uso de Animais e aprovada sob o número de protocolo 056/12. Foram utilizadas duas alturas, plantas até 1,5 m de altura e entre 1,6 – 3,0 metros, medidos a partir do ponto de inserção das folhas no solo até a curvatura da última folha.
As folhas foram submetidas ao processo de fenação por 48 horas até atingirem teor de MS médio acima de 87% e, posteriormente, trituradas em máquina forrageira em partículas de 1-2 cm. Os tratamentos químicos consistiram em amonização por ureia a 2, 4 e 6% (46, 92 e 138 g, respectivamente para o total de feno utilizado) com base na matéria seca (MS), diluídos em aproximadamente 1 L de água. A quantidade de ureia foi calculada de acordo com a quantidade de material (2,5 kg de feno em cada uma das cinco repetições), e o teor de MS deste. Após a amonização, a forragem foi acondicionada em sacos plásticos pretos, os quais foram vedados, abertos após o tratamento por 35 dias e submetidos à aeração e secagem por 48 horas para eliminação do excesso de amônia e umidade.
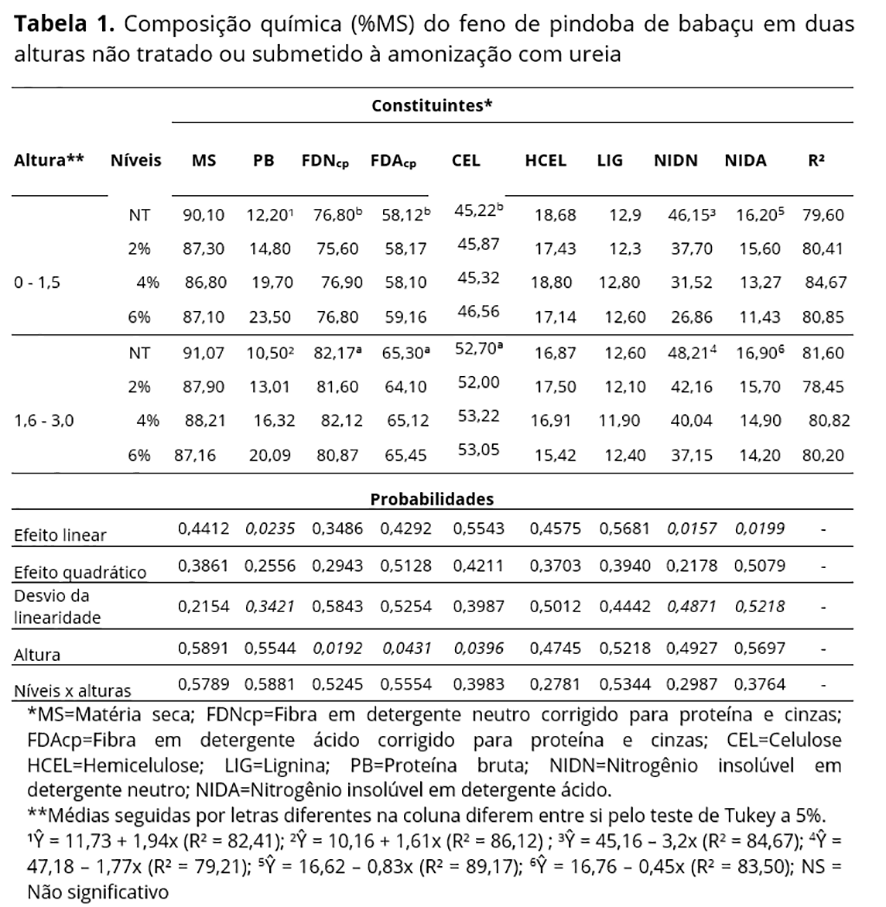
A composição química do feno não tratado e submetido à amonização com ureia foi obtida segundo metodologias da AOAC(7), para os teores de matéria seca (MS), proteína bruta (PB) e em porcentagem da PB, os teores de nitrogênio insolúvel em detergente neutro (NIDN) e nitrogênio insolúvel em detergente ácido (NIDA), segundo Licitra et al.(8). Para fibra em detergente neutro (FDN), fibra em detergente ácido (FDA) e lignina (LIG) adotou-se metodologia descrita por Van Soest et al.(9), submetidas à correção para proteína e cinzas insolúveis. Os teores de hemicelulose (HCEL) foram obtidos pela diferença entre as frações FDN e FDA e a celulose (CEL) pela diferença entre a fração FDA e a LIG.
Para avaliação da degradação ruminal, pesou-se 4 g de amostra em sacos de náilon de 12x8 cm e porosidade 50 µm de acordo com relação de 42 mg/cm² adotada por Campos et al.(10). Os sacos foram incubados no rúmen de dois bovinos adultos, fistulados, alimentados previamente com dieta composta por 80% de volumoso (capim elefante emurchecido e picado) e 20% de concentrado (milho e farelo de soja) duas vezes ao dia (8 e 16 horas), com sal mineral e água à vontade. Os tempos de incubação foram de 6, 24 e 72 horas, associados ao tempo 0 não incubado(11) utilizando-se cinco sacos como repetições em cada tratamento (4 níveis de amonização x 2 alturas) e em cada tempo, totalizando 160 sacos.
Os sacos foram incubados em ordem decrescente no tempo estipulado para cada amostra, permitindo a retirada simultânea deles após 72 horas, sendo imersos em água gelada para cessar a fermentação. A fração solúvel foi obtida por lavagem dos sacos não incubados em banho maria com revolvimento a 39 °C por uma hora, e posteriormente lavados junto aos sacos retirados do rúmen, em máquina, sob agitação e água corrente, até que ela se mostrasse límpida(12). Os sacos foram submetidos a secagem em estufa com circulação forçada de ar a 55 ºC por 72 horas para determinação da composição química.
Os parâmetros de degradação in situ (a, b e c) e a degradabilidade potencial da MS, PB, foram estimados pelo modelo exponencial por Ørskov e Mcdonald(13), expresso por: DP = a + b(1-e-ct) em que, DP = percentagem real do nutriente degradado após t horas de incubação no rúmen; a = potencial máximo de degradação do material no saco de náilon; b = fração potencialmente degradável do material que permanece no saco de náilon após o tempo zero; c = taxa de degradação da fração remanescente no saco de náilon após tempo zero; t = tempo de incubação.
A degradabilidade efetiva (DE) da MS e PB no rúmen foi estimada considerando-se a taxa de passagem 2, 5 e 8%/h para cada tratamento utilizando-se a equação proposta por Ørskov e McDonald(13): DE = a + [(b c)/(c + k)], em que, DE = degradação efetiva; a = fração solúvel, rapidamente degradada; b = fração insolúvel, lentamente degradada; c = taxa fracional de degradação de b; k = taxa de passagem.
Para estimativa dos parâmetros de degradação da FDN, adotou-se modelo de Mertens e Loften(14): Rt=B.e-ct+I, em que: Rt=fração degradada no tempo t; I = fração indegradável. Após os ajustes da equação de degradação da FDN, precedeu-se a padronização das frações, utilizando-se as equações: BP=B/(B+I)×100 e IP=I/(B+I)× 100, em que: BP = fração potencialmente degradável padronizada (%) IP = fração indegradável padronizada (%).
Adotou-se delineamento inteiramente casualizado em esquema fatorial 4x2 (quatro níveis de amonização e duas alturas) e cinco repetições por tratamento para parâmetros da composição química, enquanto que, para degradabilidade in situ da MS, PB e FDN, adotou-se delineamento em blocos (bovinos fistulados) em parcelas subdivididas com tratamentos (altura de corte x nível de ureia), representando as parcelas os tempos de incubação (0, 6, 24 e 72 horas), as subparcelas e cinco repetições por tratamento (sacos de náilon).
Os dados foram analisados quanto à normalidade e homogeneidade dos resíduos e variâncias e realizado teste de regressão para determinar os efeitos das doses de ureia sobre os componentes químicos, bem como para verificar interação entre alturas de corte e doses de ureia e o desvio da linearidade pelo procedimento PROC GLM do SAS (2000). Para altura de corte do feno não tratado, as médias foram comparadas pelo teste de Tukey a 5%. Quanto à avaliação dos parâmetros de degradação in situ, foram utilizados procedimentos para modelos não lineares (PROC NLIN) do pacote estatístico SAS (2000), estabelecido com base nos ajustes das frações e na curva de degradação obtidas pelos modelos propostos para degradação da MS, PB e FDN.
Resultados e discussão
A amonização contribuiu para incremento (P<0,05) dos teores de PB nas alturas de corte, com elevação média de 20% no teor proteico a cada aumento na doses de ureia, em virtude da incorporação de nitrogênio não proteico (NNP) ao material, e para menores (P<0,05) teores de NIDN e NIDA (Tabela 1), efeitos constatados por Bezerra et al.(15) para feno de capim Buffel e Andrade et al.(16), com incrementos de 17 e 22% na PB com adição de 8% de ureia, respectivamente. Não houve desvio da linearidade para os compostos nitrogenados, nem interação significativa para estes entre as alturas de corte e doses de ureia.
A elevação na proporção de NNP no feno pode contribuir para melhor degradação dos constituintes fibrosos, pois baixos teores de N insolúvel indicam maior disponibilidade desse nutriente no ambiente ruminal, utilizado preferencialmente por microrganismos celulolíticos para fixação em esqueletos de carbono e formação de proteína microbiana(3,19).
Os teores de PB obtidos nessa pesquisa para o feno da pindoba de babaçu nas duas alturas de corte foram superiores aos obtidos para gramíneas forrageiras comumente utilizadas em dietas para ruminantes por Martins-Costa et al.(17) e Velásquez et al.(18) para capim elefante (9,70%) e feno de capim Tânzania (9,75%) respectivamente, e acima do mínimo de 6-8% na MS, para manutenção de 8mg de N-NH3.dL-1 de liquido ruminal, recomendado para eficiente degradação ruminal e incremento na produção de proteína microbiana(19), o que denota a possibilidade de seu uso como fonte de volumoso, quanto aos teores desse constituinte.
Os teores de nitrogênio insolúvel em detergente ácido (NIDA) reduziram (P<0,05) de 0,83 e 0,45% com o aumento da porcentagem de ureia na MS, respectivamente para altura de até 1,5 e de 1,6 – 3,0 m, sendo inferiores a proporção de 20% do N total, considerado por Van Soest(9) limitante à degradação da forragem, por redução do N para ação microbiana. Essa fração nitrogenada corresponde à proteína associada à lignina, complexos taninoproteína e produtos oriundos da reação de Maillard, altamente resistentes às enzimas microbianas e indigestíveis ao longo do trato gastrintestinal(20).
A amonização com ureia não influenciou os teores de FDNcp e FDAcp do feno de pindoba de babaçu, sem efeito linear ou desvio na linearidade, todavia houve aumento (P<0,05) dessas frações na maior altura da forrageira (1,6 – 3,0 m), com elevação de 4,07% para FDNcp e 7,33% para FDAcp no feno não tratado. Esses teores estão acima dos obtidos por Garcez et al.(4) ao avaliarem o feno dos folíolos de pindoba de babaçu, com médias de 67,6% e 49,6% para FDN e FDA, respectivamente, o que está associado às partes da planta utilizada; nesta pesquisa avaliou-se a folha completa, o que elevou os teores de constituintes fibrosos por incorporação oriunda do colmo central.
O aumento (P<0,05) no percentual de FDN e FDA com o avanço na idade indica redução na proporção de nutrientes da fração fibrosa passíveis de serem digeridos pelo animal refletindo diretamente no consumo de MS e metabolismo microbiano, fato atribuído ao espessamento e lignificação da parede celular(21). Não houve interação entre as doses de ureia e as alturas do feno de pindoba de babaçu para as frações fibrosas, hemicelulose (HCEL), celulose (CEL) e Lignina (LIG) (P>0,05), o que aponta efeito semelhante desse tratamento alcalino sobre os constituintes da parede celular, sem variações mesmo com o avançar da idade da planta.
Houve aumento na degradação potencial da MS com a elevação das doses de ureia, com destaque para o tratamento com 4% e altura de 1,5m (Tabela 2). No entanto os valores para esse tratamento quanto à fração b (17,22%) e à degradação potencial (DP) (28,45%) foram inferiores aos obtidos por Garcez et al.(4) para feno de folíolos de pindoba de babaçu (18,91 e 32,92%), associados aos maiores de fibra obtidos nessa pesquisa. A taxa de degradação (c) aumentou em média 0,26 (1,5 m) e 0,28%/h (3,0 m) a partir do tratamento com 4% de ureia; no entanto, reduziu com a maior altura da forrageira, refletindo na redução do valor nutritivo e disponibilidade de nutrientes da planta.
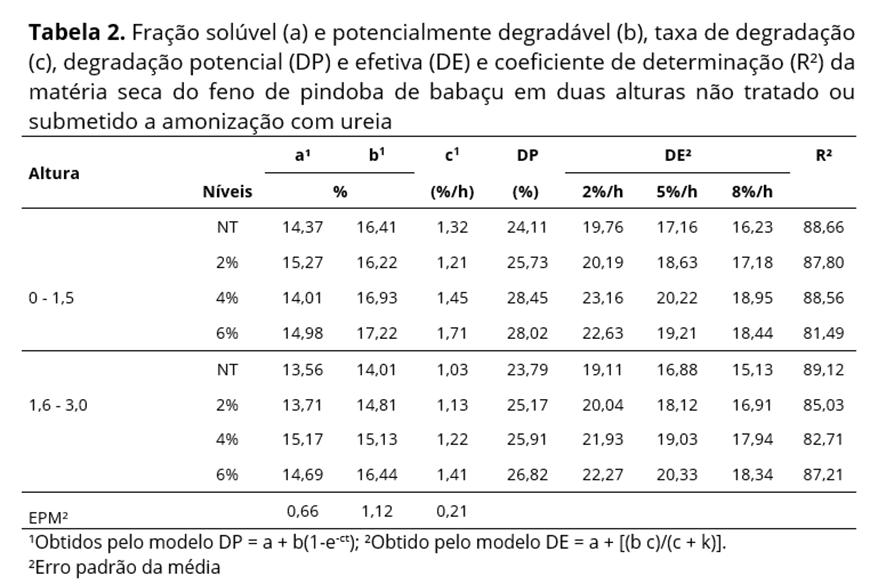
As reduções da DP e o menor efeito da amonização com a maturidade da pindoba de babaçu associam-se ao aumento da proporção de colmo nas folhas verificada quando do corte e fenação da forrageira. O colmo tem em sua composição tecidos formados por células densas, espessas e lignificadas, o que reduz a disponibilidade de celulose e hemicelulose para a degradação ruminal, com consequente déficit de energia oriunda da glicose presente nesses polissacarídeos(22). O elevado teor de FDN (82,17%) e elevadas proporções de lignina (acima de 12%) estão associadas às menores taxas de degradação da MS obtidas para o feno de pindoba de babaçu nesse estudo.
A fração solúvel (a) da PB foi elevada conforme o aumento das doses de ureia, relacionado à maior solubilidade desse composto nitrogenado (Tabela 3). A DP apresentou maiores valores no tratamento com 4% de ureia em ambas as alturas, com incrementos acima de 10%, porém inferiores aos obtidos por Moreira Filho et al.(6) para feno do restolho da cultura do milho amonizado com 3% de ureia.
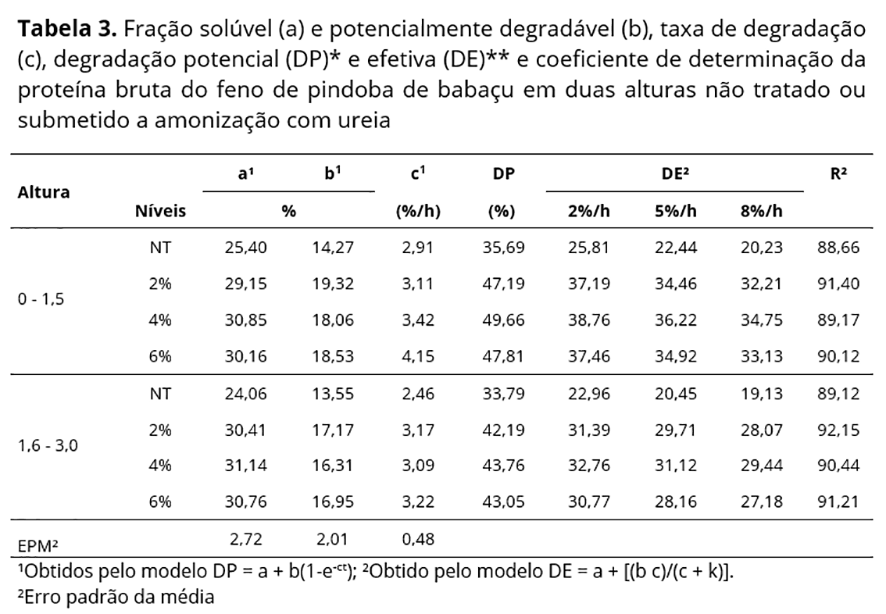
A degradação potencial da proteína bruta influencia o aproveitamento ruminal dos demais componentes nutricionais, em que as menores taxas reduzem a quantidade de N disponível para o metabolismo, e consequentemente, na síntese de proteína microbiana(22). Os valores de DP obtidos nesta pesquisa foram inferiores aos obtidos para feno de capim tifton em idade avançada (74,12%) por Jobim et al.(23) e para capim elefante com 110 dias de corte (64,27%) por Rêgo et al.(24), o que denota menor disponibilidade de N mesmo com teores reduzidos de nitrogênio insolúvel (NIDA) (Tabela 1), com elevação dessa fração apenas com a inclusão de NNP oriundo da ureia.
A redução na DP da PB com elevação da altura da pindoba de babaçu segue padrões de crescimento de gramíneas tropicais, devido à complexação com carboidratos da parede celular, formando ligações do tipo éster com compostos fenólicos presentes na fração FDA e passando a fazer parte da fração insolúvel do alimento, interferindo na colonização bacteriana, com menor degradação dos demais componentes nutritivos da dieta(18).
A amonização com ureia reduziu o tempo de colonização (lag) e elevou a fração potencialmente degradável (Bp) da FDN com maiores valores para doses acima de 2% na MS na menor altura (Tabela 4). A redução no lag com adição de ureia ocorre em virtude da melhor eficiência na aderência microbiana às frações fibrosas, com o incremento de NNP disponível no rúmen, maximizando o crescimento de bactérias degradadoras de fibra, que se utilizam desse composto para fixação de N em seus aminoácidos durante a síntese de proteína microbiana(25).
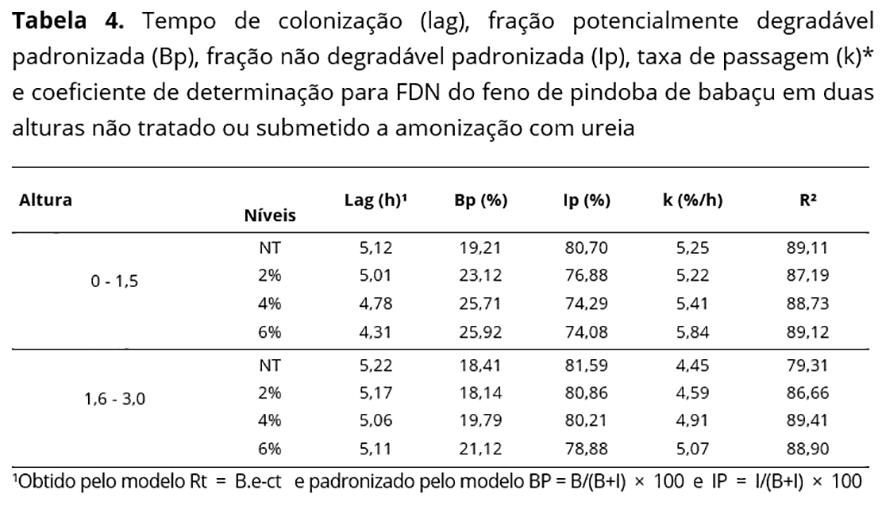
Os valores de Bp da FDN são considerados baixos quando comparados a volumosos de nativos avaliados por Muniz et al.(26) obtendo valores de 31% para feno de mata-pasto (Senna obtusifolia L.) e 28,91% para mororó (Bauhinia cheilanta). O aumento dessa fração com a amonização é atribuído à ação da amônia na quebra das ligações entre a hemicelulose e a lignina e aumento no potencial de solubilidade da fração fibrosa, o que justifica maiores valores na menor altura, que tem menos ligações fenólicas e maior disponibilidade de carboidratos da parede celular.
A fração indegradável da FDN (Ip) correspondeu à maior parte da fração fibrosa, o que pode ser o principal determinante da baixa degradação da MS, em virtude de a FDN corresponder a mais que 70% desse constituinte. O aumento dessa fração com o decorrer da idade da forrageira é esperado, uma vez que espécies de regiões semiáridas atingem maturidade fenológica precoce com aumento de estruturas de sustentação representadas pelos carboidratos fibrosos e lignina(26), sendo superiores aos obtidos por Garcez et al.(4) com 71,70% para feno de folíolos de pindoba de babaçu amonizado com 6% de ureia, associado à inclusão do colmo central mais fibroso quando do preparo do feno nesta pesquisa.
Em termos nutricionais, deve-se considerar que todos os eventos de ação microbiana nas frações potencialmente degradáveis e indegradáveis da FDN ocorrem em escalas de tempo finitas. Assim, a forma de ação sobre o substrato depende do tempo no qual a interação enzima x substrato ocorre, ou seja, da efetividade do processo de degradação(27). Nesse sentido, a efetividade do processo de degradação da FDN no ambiente ruminal é diretamente representada pela estimativa da fração efetivamente degradada da FDN, a qual associa a velocidade de ação enzimática sobre o substrato e o tempo disponível para que estes sistemas executem tais ações.
Conclusão
A amonização com ureia eleva os teores de PB do feno de pindoba de babaçu nas duas alturas devido à inclusão de nitrogênio não proteico, porém não interfere nos teores de FDNcp e FDAcp, com elevação destes com o avançar da altura da planta. O feno de pindoba de babaçu apresenta melhor degradação potencial e efetiva da MS, PB e FDN quando obtido em altura até 1,5 m e submetido a amonização com doses acima de 4% de ureia, sendo indicado essa altura e esse nível de amonização para se obter um feno de melhor qualidade.
Referências