Com o objetivo de estudar os
limitantes da degradação da fibra da cana-de-açúcar para utilizá-la
como alimento para bovinos, fracionaram-se os componentes fibrosos da
cana-de-açúcar (variedade Mex69-290) em fibra em detergente neutro
(FDN) e em fibra em detergente ácido (FDA). A cana integral e as fibras
foram incubadas com bactérias ruminais, na presença ou ausência de
enzimas fibrolíticas (Fibrozyme, Alltech Inc.). Determinaram-se o
crescimento microbiano e as bactérias totais, assim como a taxa de
crescimento, a taxa de geração, o tempo de colonização e a
degradabilidade. Os resultados foram analisados em um delineamento de
blocos ao acaso, com arranjo fatorial 2 x 3, com duas doses de enzima e
três substratos, utilizando quatro repetições de incubação. Observou-se
o maior crescimento microbiano para cana integral e os menores para
suas fibras. A degradabilidade in vitro foi de 0,701, 0,392 e 0,191,
respectivamente para cana integral, FDN e FDA (P<0,01). A fração FDA
com a adição das enzimas fibrolíticas teve sua degradabilidade
aumentada de 0,387 para 0,425 (P<0,01). O crescimento microbiano foi
limitado nas frações fibrosas, possivelmente pelo grau de lignificação
das paredes celulares. A maior degradabilidade in vitro das frações
fibrosas está associada à presença de açúcares solúveis. A adição de
enzimas fibrolíticas aumentou o crescimento microbiano máximo e a
digestibilidade da FDA, indicando que estas podem ser um potencial
aditivo para melhorar o aproveitamento de dietas com cana-de-açúcar.
PALAVRAS-CHAVES: Crescimento microbiano, degradabilidade in vitro, parede celular.
Aiming to study the limiting factors
for degradation of sugarcane fiber to be used as cattle feed, sugarcane
fibrous components were fractioned in neutral- and acid-detergent
fibers (respectively, NDF and ADF). Whole sugarcane and its fibers were
incubated with rumen bacteria, in presence or absence of fibrolytic
enzymes (Fibrozyme, Alltech Inc.). Microbial growth and total bacteria
count were determined, and the growth rate, generation rate, lag time,
and degradability were also determined. Results were analyzed in
randomized block design, with a 2x3 factorial arrangement, with two
doses of enzymes, and three substrates, using four replications for
incubation. The highest microbial growth was observed for whole
sugarcane, and the lowest for its fibers. The in vitro degradability
was 0.701, 0.392 and 0.191, respectively for whole sugarcane, NDF and
ADF (P<0.01). ADF fraction with the addition of fibrolytic enzyme
had its degradability increased from 0.387 to 0.425 (P<0.01).
Microbial growth was limited in fibrous fractions, possibly due to
lignification grade of cell walls. The highest in vitro degradability
of fibrous fractions is related to the presence of soluble sugars.
Addition of fibrolytic enzymes increased the maximum microbial growth
and ADF degradability, indicating that it could be a potential
addictive to enhance diets containing sugarcane.
KEYWORDS: Cell wall, in vitro degradability, microbial growth.
Os principais limitantes da utilização da cana-de-açúcar (
Saccharum officinarum
L.) na alimentação de ruminantes são a baixa digestibilidade de sua
parede celular, o elevado teor de açúcares e a estrutura das moléculas
das paredes celulares (LOPEZ
et al., 2003; ARANDA
et al., 2004). GÓMEZ-VAZQUEZ
et al.
(2003) notaram uma resposta linear na digestibilidade da fibra em
detergente neutro (FDN) e no ganho de peso dos novilhos alimentados com
pasto estrela e cana-de-açúcar, ao receber doses crescentes de uma
enzima fibrolítica exógena. O objetivo deste trabalho foi estudar o
efeito da adição de enzimas fibrolíticas exógenas sobre o crescimento
microbiano e sobre a fermentação ruminal da cana-de-açúcar, sob a
hipótese de que as enzimas fibrolíticas podem incrementar a
digestibilidade das paredes celulares da cana-de-açúcar, assim como
melhorar a eficiência da fermentação ruminal e estimular o crescimento
microbiano.
MATERIAL E MÉTODOS
Foram realizadas incubações in vitro de cana-de-açúcar integral, variedade Mex69-290 (
Quadro 1), assim como de suas frações fibrosas extraídas com detergentes neutro e ácido (respectivamente FDN e FDA) (VAN SOEST
et al.,
1991) com ou sem a adição da enzima fibrolítica Fibrozyme© (Alltech
Inc.), na dose de 100 mg/g de substrato incubado (PINOS, 1999).
Calcularam-se a taxa de crescimento com base na regressão do logaritmo
natural da concentração de bactérias em função do tempo e a taxa da
geração mediante a relação 0,693/k, sendo k a taxa específica de
crescimento ou o coeficiente de manutenção da relação inversa da
concentração das bactérias, no tempo zero (PIRT, 1982). O tempo de
colonização (fase lag) foi estimado como inverso dos valores do
crescimento por extrapolação ao tempo zero (ZWIETERING
et al., 1991). Estimaram-se os parâmetros por regressão (DRAPPER & SMITH, 1981).
Em erlenmeyers de capacidade de 250 mL, foram colocados 150 mL de meio anaeróbio (
Quadro 2),
1 mL de líquido ruminal e 100 mg de substrato incubados em saco de
nylon, medindo 4 x 5 cm, por 24 horas à temperatura constante de 39°C.
Para isso, utilizaram-se dez réplicas por amostra, com e sem a adição
de enzima fibrolítica. Ao final da incubação, os sacos foram secos a
65°C, até peso constante. Considerou-se a diferença entre os pesos
inicial e final das amostras como material degradado e calculou-se a
degradabilidade como a razão entre o material degradado e o peso
inicial.
Seguindo a mesma metodologia, como descrito, incubaram-se as amostras
com e sem enzima fibrolítica, com três réplicas, repetidas em cinco
ensaios. O crescimento microbiano foi determinado em intervalos de
trinta minutos nas primeiras dez horas, com uma leitura final às 25
horas, utilizando espectrofotômetro (Spectronic 20, Bausch and Lomb),
ajustado para 600 nm, segundo RUSSEL & DOMBROWSKI (1980).
Empregou-se a contagem total de bactérias, para transformar o valor de
densidade óptica em concentração de bactérias, visando caracterizar as
curvas de crescimento (MIRANDA, 1998).
Os resultados foram analisados de acordo com um delineamento de blocos
ao acaso (STEEL & TORRIE, 1980), com arranjo fatorial 2 x 3, com
dois teores de enzima e três substratos (cana integral e suas frações –
FDN e FDA). Realizaram-se quatro repetições de incubação (CLARY
et al.,
1988). Usou-se o seguinte modelo estatístico: Yijk = µ + Bj + Si + Ck+
Si*Ck + Bj*Si*Ck + εijk, sendo Yijk a variável resposta; µ, a média
geral; Bj, efeito do bloco; Si, efeito do substrato; Ck, efeito da
adição de enzima; Si*Ck e Bj*Si*Ck, interações; e εijk, o erro residual
do modelo. Empregou-se o procedimento GLM do SAS (1985),
considerando-se a interação blocos x tratamentos como o erro
experimental. As médias foram comparadas pelo teste de Tukey (STEEL
& TORRIE, 1980).
RESULTADOS E DISCUSSÃO
Na
Tabela 1 e na
Figura 1, é apresentado o crescimento das bactérias ruminais de acordo com o substrato incubado.
Apesar de haver uma tendência de maior crescimento microbiano na cana
integral, as diferenças estatísticas com respeito ao substrato foram
observadas a partir das 3,5 horas. Nas primeiras horas de incubação,
não houve diferenças significativas, possivelmente em virtude da grande
variabilidade da absorbância, coincidindo com as observações de MERTENS
(1993), que apontam que os trabalhos in vitro, em sua primeira etapa,
apresentam alta variabilidade. O maior crescimento microbiano na cana
integral se deve à presença de açúcares solúveis de fácil degradação
(BANDA & VALDEZ, 1976), e o menor crescimento em FDA se associa ao
alto teor de lignina (AMJED
et al., 1992).
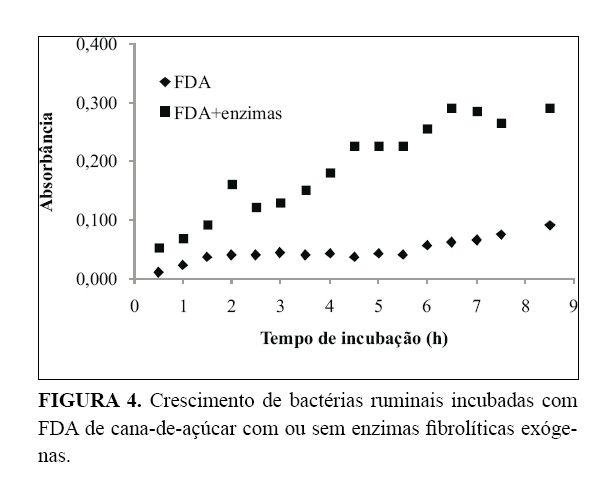
A resposta do crescimento microbiano mediante a adição de enzimas é apresentada nas
Figuras 2,
3 e
4, sendo uma para cada substrato. Na
Tabela 2,
os principais efeitos são apresentados, e nela pode-se observar o maior
crescimento microbiano em resposta à adição da enzima. Há uma resposta
positiva à adição de enzimas fibrolíticas, indicando que há aumento da
disponibilidade de metabólitos para o crescimento microbiano. A maior
diferença foi observada na fração FDA (
Figura 4),
a qual poderia estar associada à maior degradabilidade devida à ação
enzimática sobre as ligações ligno-celulósicas e possivelmente a outros
efeitos indiretos. É possível que a adição de enzimas exógenas possa
incrementar a fração potencialmente degradável da celulose contida na
FDA (AKIN, 1986).
Na
Tabela 3, são apresentados os parâmetros de crescimento microbiano na cana-de-açúcar. Apesar das diferenças apresentadas na
Tabela 2,
as diferenças estatísticas não são detectadas para os parâmetros do
crescimento microbiano na cana-de-açúcar e em suas frações fibrosas.
Esse problema foi detectado em análise de cinética de primeira ordem.
MENDOZA
et al. (1995) sugerem
que os dados sejam analisados por tempo de incubação, dado que podem
existir diferenças em alguns tempos de incubação, e a linearização com
logaritmo natural altera o erro residual do modelo, não permitindo
detectar estatisticamente algumas diferenças biologicamente importantes.
As taxas de crescimento e as taxas de geração foram similares para cana integral e para as frações FDN e FDA (
Tabela 3).
As taxas de crescimento observadas neste experimento são menores que as
reportadas para cultivos mistos em substratos ligno-celulósicos
(1,13/h) como os restos da cultura de milho (MIRANDA
et al., 1999). Os microrganismos celulolíticos
F. succinogenes, R. flavefaciens e
R. albus
têm a capacidade de degradar a celulose a uma taxa entre 0,05 e 0,10/h
(WEIMER, 1996). É possível inferir que a limitada taxa de crescimento
na cana-de-açúcar é influenciada pela lignificação da fibra (
Quadro 1)
e pode ser o principal limitante para o aproveitamento da
cana-de-açúcar, tornando necessária a busca de alternativas para
aumentar a digestibilidade destas frações com tratamentos físicos,
químicos, enzimáticos e biológicos. O tempo de colonização tendeu a ser
maior na cana integral (
Tabela 3).
Tem sido reportados valores de 2,5 a 3,0 h de tempo de colonização em
incubações com restos da cultura de milho com 70% de FDN (MIRANDA
et al.,
1999), os quais são menores que os observados neste estudo. A fase lag
na FDA da cana-de-açúcar confirma a importância da lignina como
limitante na digestão desse alimento.
A degradabilidade in vitro da cana-de-açúcar e suas frações fibrosas
refletem a atividade microbiana e apontam os limitantes inerentes das
paredes celulares para a digestão animal. A maior degradabilidade da
cana-de-açúcar se deve à concentração de açúcares solúveis no conteúdo
celular (BANDA & VALDEZ, 1976; AROEIRA et al., 1993a; AROEIRA et
al., 1993b).
Apenas se observaram diferenças estatísticas na degradabilidade, sendo
maior a da cana-de-açúcar, seguida por FDN e FDA. A degradabilidade
in vitro
da FDA observada neste estudo está dentro do intervalo de valores
reportados (PATE, 1977) e coincide com a FDN potencialmente degradável
reportada para subprodutos da cana (AMJED et al., 1992) associada à
relação linear negativa entre a degradabilidade in vitro da FDN e a
proporção FDN:lignina (PATE, 1977). A menor degradabilidade da FDA
confirma a importância da lignina como limitante no aproveitamento da
cana pelos microrganismos ruminais.
Na
Tabela 4,
são apresentados os principais efeitos das enzimas fibrolíticas nos
parâmetros de crescimento microbiano e na degradabilidade in vitro. A
adição da enzima aumentou a degradabilidade dos substratos em 3,81
unidades percentuais, o que pode ser a explicação do maior crescimento
microbiano máximo, sem afetar outros parâmetros do crescimento. Tem
sido reportados aumentos na degradabilidade in vitro da MS, da FDN e da
FDA de gramíneas e leguminosas com a adição da enzima Fibrozyme© (FENG
et al., 1996; TRICARICO et al., 1998; PINOS et al., 2001; PINOS et al.,
2002a; PINOS et al., 2002b) e mesmo na digestibilidade in vivo (KRAUSE
et al., 1988; BEAUCHEMIN et al., 1998). No entanto, os resultados têm
sido muito variáveis, possivelmente em virtude da razão
enzima:substrato e da degradação da enzima por proteases ruminais
(HARRIS, 1998).
As enzimas utilizadas são uma combinação de celulases e hemicelulases
produzidas por fungos, protegidas por glicosilação, e estima-se que
podem permanecer ativas por volta de doze horas no rúmen (HARRIS, 1998;
LYONS, 1998). PINOS (1999) estudou a composição e a degradação ruminal
in vitro da enzima Fibrozyme e reportou que o complexo tem 93,6% de MS,
45,8% de FDN, 32,5% de FDA e 9,5% de cinzas, encontrando um tempo médio
de degradação do complexo de 57 horas, com uma maior liberação de
nitrogênio amoniacal depois de 24 horas.
O uso de enzimas microbianas celulolíticas exógenas para incrementar a
digestibilidade da fibra da cana-de-açúcar é uma alternativa viável,
como mostram os resultados de GÓMEZ-VÁSQUEZ et al. (2003). Também
alguns estudos com gramíneas e leguminosas de clima temperado têm
mostrado que, com a aplicação de enzimas fibrolíticas, podem ser
melhorados a digestibilidade e o crescimento de novilhos em cerca de
30% (BEAUCHEMIN et al., 1996) e a produção de leite em até 10%
(BEAUCHMIN et al., 1998; KUNG et al., 1998; YANG et al., 1998).
Juntamente com o conhecimento das estruturas da parede celular da
cana-de-açúcar e das enzimas industriais, é possível desenvolver novas
alternativas de tratamentos para melhorar o aproveitamento da celulose
e da hemicelulose pelos ruminantes.
CONCLUSÕES
O crescimento microbiano a partir da degradação da cana-de-açúcar é
limitado pelas frações fibrosas, possivelmente pelo grau de
lignificação das paredes celulares. A maior degradação in vitro da
cana-de-açúcar em relação às suas frações fibrosas está associada à
presença de açúcares solúveis.
A adição de enzimas fibrolíticas aumentou o crescimento bacteriano
máximo e a degradabilidade da FDA, podendo-se concluir que é possível
considerar as enzimas fibrolíticas como um aditivo potencial para
melhorar o aproveitamento de dietas com cana-de-açúcar.
REFERÊNCIAS
Amjed, M.; Jung,
H. G.; Donker, J. D. Effect of
alkali hydrogen peroxide treatment on cell wall composition and digestion
kinetics of sugarcane residues and wheat straw. Journal
of Animal Science, v. 70, p. 2877-2884, 1992.
ARANDA, E. M.; RUIZ, P.; MENDOZA, G. D.; MARCOFF, C. F.; RAMOS, J.
A.; Y ELÍAS A. Cambios en la digestión
de tres variedades de caña de azúcar y sus fracciones de fibra. Revista Cubana de Ciencia Agrícola,
Tomo 38, n. 2, p. 137-144, 2004.
Aroeira, R. S.; Figueira, D. G.; Rodriguez, N. M.;
Sampaio, I. B. M.; Lopes, F. C.; Torres, e. M. P. Degradabilidade in
situ dos nutrientes da cana-de-açúcar e do farelo de algodão em bovinos
alimentados com farelo de algodão e cana-de-açúcar de três níveis de uréia. Arquivo Brasileira de Medicina Veterinária
Zootecnia, v. 45, p. 221-233, 1993b.
Aroeira, R. S.;
Lizieire, R. S.; Matos, L. L.; Figueira, D. G. Rumen degradability and rate of passage of sugar
cane + urea based diets, supplemented with cottonseed or rice meals in Holstein
x Zebu steers. Journal of Animal Science, v. 71 (supp.1), p. 273,
1993a.
Banda, M.; Valdez, R. E. Efecto del estado de madurez sobre el valor nutritivo
de la caña de azúcar. Produção Animal nos Trópicos, v. 1, p. 96-99,
1976.
Beauchemin, K.
A.; Rode, L. M.; Sewalt, V. J. H. Fibrolitic enzimes increase fiber
digestibility and growth rate steers fed dry forages. Canadian Journal of
Animal Science, v. 75, p. 641-644, 1996.
Beauchemin, K.
A.; Yang, W. Z.; Rode, L. M.
Effects of fibrolytic enzyme additives on extent of digestion and milk
production of lactating cows. Journal of Animal Science, v. 76 (Suppl.
1), p. 358, 1998.
Clary, W. P.;
Welch, B. L.; Booth, G. D. In vitro digestion
experiments: importance of variation between inocula donors. Journal of
Wildlife Management, v. 52, p. 358-361, 1988.
Drapper, N.;
Smith, H.
Applied regression analysis. New York: John Wiley & Sons,
1981. 709 p.
Feng, P. C.;
Hunt, W.; Pitchard, G. T.; Julien, d. W. E. Effect of enzyme preparations on in situ and in
vitro digestive characteristics of mature cool-season grass forage in beef
steers. Journal of Animal Science, v. 74, p. 1349-1357, 1996.
forages. Journal
of Animal Science, v. 63, p. 962-977, 1986.
Gomez-Vazquez, A.; Pérez, J.; Mendoza, G. D.; Aranda,
E.; Hernández, A. Fibrolytic exogenous enzymes improve perfomance in
steers fed sugar cane stargrass. Livestock Production Science, v. 82, p.
249-254, 2003.
Harris, B. The emerging role of enzymes in ruminant
diets: at long last, a breakthrough. Udder Information. Dr. Harri´s Guide to
Maximizing Dairy Performance. 1998. Disponível em:
<http://www.alltech-bio.com/udder98.htm.pp1-13.> Acesso em: 12 jun. 2006.
Krause, M.;
Beauchemin, K. A.; Rode, L. M.; Farr, B. I.; Norgaard, P. Fibrolytic enzyme treatment of barley grain
and source of forage in high-grain diets fed to growing cattle. Journal of
Animal Science, v. 76, p. 2912-2920, 1988.
Kung, L.;
Treacher, R. J.; Cohen, M. A. Enzyme-treated
forages for lactating cows. Journal of Animal Science, v. 76 (Suppl. 1),
p. 196, 1998.
LÓPEZ,
I.; ARANDA, I. E. M.; RAMOS, J. J. A; MENDOZA, M. G. D. Evaluación nutricional
de ocho variedades de caña de azúcar con potencial forrajero. Revista
Cubana Ciencia Agricola, v. 37, p. 381. 2003.
Lyons, T. P. The consumer is king: where sill it all
end for the deed industry. In: Lyons, T.
P.; Jacques, K. A. (Eds.).
Biotechnology in the feed industry. Proceedings of the Fourteenth Annual
Symposium. Loughborough: Nottingham University Press, 1998. p. 3-29.
Mendoza, M. G. D.; Ricalde, V. R.; Esparza, H. B.;
Velazquez, y. L. T. Nota: Efecto de dos
cultivos de Saccharomyces cerevisiae en la degradacion ruminal de la
fibra neutro detergente de paja de trigo. Investigación Agricola, Producción y Sanidade Animales, v. 10, p. 33-38, 1995.
MERTENS, D. M. Kinetics of cell wall
digestion and passage in ruminants. In: JUNG, H. G.; BUXTON, D. R.; HATFIELD,
R. D.; RALPH, J. ASA-CSSA-SSSA, 677S. Forage cell wall structure and
digestibility. Madison, USA: Segoe RD. 1993. p. 535-571.
MIRANDA,
R. L. A.; COBOS, M. A. P.; MENDOZA, M.
G. D.; GONZÁLEZ, M. S. S.; GARCÍA, B. C. M. Degradación in vitro de
rastrojo de maíz con cultivos mixtos de bacterias ruminales. Agrociencia, v. 33, n. 2, p. 133-139. 1999.
Pate, F. M. Nutritive value of sugar cane at
different stages of maturity. Tropical Animal Production, v.
2, p. 108-109, 1977.
PINOS-RODRÍGUEZ,
J. M.; GONZÁLEZ-MUÑOZ, S. S.;
MENDOZA-MARTÍNEZ, G. D.; BÁRCENA-GAMA
R.; COBOS-PERALTA. M. Efecto de enzimas fibrolíticas glucosiladas en la
digestibilidad in vitro de MS y MO de alfalfa (Medicago sativa) y
ballico (Lolium perenne). Revista Científica, FCV-LUZ v. 11, n.
6, p. 505-509. 2001.
PINOS-RODRÍGUEZ,
J. M.; GONZÁLEZ-MUÑOZ, S. S; MENDOZA-MARTÍNEZ, G. D.; BÁRCENA-GAMA R.; COBOS-PERALTA, M. Efecto de enzimas fibrolíticas exógenas en la
digestibilidad in vitro de la pared celular del heno de alfalfa (Medicago
sativa) o de ballico (Lolium perenne). Interciencia, v. 27, n. 1, p. 28-32. 2002a.
PINOS-RODRÍGUEZ, J. M.; GONZÁLEZ-MUÑOZ,
S. S.; G.D. MENDOZA, G. D.; R. BÁRCENA, R.; COBOS, M. A.; HERNÁNDEZ, A.;
ORTEGA, M. E. Effect of exogenous fibrolytic enzyme on ruminal fermentation and
digestibility of alfalfa and rye-grass hay fed to lambs. Journal of Animal
Science, v. 80, n. 11, p. 3016-3020. 2002b.
Pirt, S. J. Maintenance energy: a general model for
energy-limited and energy-sufficient growth. Archives of
Microbiology, v. 133, p. 300-302, 1982.
Russell, J. B.; Dombrowski, D. B. Effect of pH on the efficiency
of growth by pure cultures of rumen bacteria in continuous culture. Applied
Environmental Microbiology, v. 39, p. 604-610, 1980.
Statistical
Analysis System. S.A.S.
User’s Guide: statistics version. 5. ed. Cary: SAS Institute Inc., 1985.
Steel, R. G. D.;
Torrie, J. H. Principles and procedures of
statistics: a biometrical approach. 2. ed. New York: McGraw-Hill Book Co.,
1980. 633 p.
Tricarico, M.;
Dawson, K. A.; Newman, K. E.
Effects of an exogenous microbial enzyme preparation (Fibrozyme) on ruminal
digestion of fescue hay. Journal of Animal Science, v. 76 (Suppl. 1), p.
289, 1998.
Van Soest, P. J.;
Robertson, J. B.; Lewis, B. A.
Symposium: carbohydrate methodology, metabolism, and nutritional implications
in dairy cattle. Journal of Dairy Science, v. 74, p. 3583-3597, 1991.
Weimer, P. J.
Why don’t ruminal bacteria digest cellulosa faster. Journal of Dairy
Science, v. 79, p. 1496-1502, 1996.
Yang, W. Z.;
Rode, L. M.; Beauchemin, K. A.
Effects of fibrolitic enzyme additives on milk production of dairy cows. Journal
of Animal Science, v. 76 (Suppl. 1), p. 320, 1998.
Zwietering, M.
H.; de Koos, J. T.; Hasenack, B. E.; de Wit, J. C.; Van´T Riet, K. Modeling of bacterial growth as a
function of temperature. Applied Environmental Microbiology, v.
7, p. 1094-1101, 1991.
Protocolado
em: 17 set. 2008. Aceito em: 20 maio 2010.
{kind=link}