DOI:
10.1590/1809-6891v20e-44460
ZOOTECNIA
COMO A IDADE DO PERFILHO E A ADUBAÇÃO NITROGENADA
MODIFICAM AS CARACTERÍSTICAS ESTRUTURAIS DO CAPIM-MARANDU DIFERIDO?
HOW DO AGE TILLER AND NITROGEN FERTILIZING MODIFY THE STRUCTURAL
CHARACTERISTICS OF DEFERRED MARANDU PALISADE GRASS?
Angélica Nunes de Carvalho1 ORCID http://orcid.org/0000-0002-8233-5909
Lucas Coelho Alves1 http://orcid.org/0000-0001-6744-4018
Manoel Eduardo Rozalino Santos1* http://orcid.org/0000-0002-3668-4518
Gabriel de Oliveira Rocha1 ORCID http://orcid.org/0000-0002-5557-2458
Pedro Henrique Marçal Rodrigues1 ORCID http://orcid.org/0000-0003-1872-4579
Bruno Humberto Rezende Carvalho1 ORCID http://orcid.org/0000-0003-1844-0699
1Universidade Federal de Uberlândia, Uberlândia,
MG, Brasil.
*Autor para correspondência - manoel.rozalino@ufu.br
Resumo
Este estudo avaliou as características estruturais de perfilhos, com
diferentes idades, do capim-marandu (Urochloa brizantha cv.
Marandu) diferido e adubado. Foram avaliadas duas doses de nitrogênio
(baixa, 50 kg ha-1 e alta, 200 kg ha-1), três categorias de idades de
perfilhos (jovem, <2 meses; maduro, 2 a 4 meses; e velho, > 4
meses;), além de dois períodos (início, 1 a 45 dias e fim, 46 a 90 dias)
durante o diferimento. O delineamento experimental foi inteiramente
casualizado, em esquema de parcela sub-subdividida, com quatro repetições.
De modo geral, o perfilho jovem apresentou melhor composição morfológica
(40,0% de lâmina foliar viva, P=0,0074; 39,2% de colmo vivo, P=0,0208; e
20,8% de lâmina foliar morta, P=0,0155), enquanto que o perfilho velho
teve pior morfologia (7,3% de lâmina foliar viva; 54,8% de colmo vivo; e
38,0% de lâmina foliar morta) ao término do período de diferimento. A
percentagem de perfilho velho foi maior (P=0,0019) no fim (54,0%) do que
no início (43,4%) do período de diferimento. No início do diferimento o
dossel adubado com alta dose de N apresentou (P=0,0447) maior percentual
de perfilho jovem (36,8%) do que aquele sob baixa dose de N (28,6%). No
fim do período de diferimento, tanto a alta como a baixa dose de
nitrogênio resultaram (P=0,0276) em valores similares de perfilho jovem no
dossel diferido (24,3%, em média). A maior dose de N, parcelada antes do
período de diferimento, aumenta o percentual de perfilho jovem durante a
fase inicial do diferimento. O perfilho jovem tem melhores características
estruturais do que os perfilhos maduros e velhos.
Palavras-chave: área foliar; composição morfológica;
número de perfilho; Urochloa brizantha.
Abstract
The structural characteristics of tillers with different ages of deferred
and fertilized marandu palisade grass (Urochloa brizantha cv.
Marandu) were evaluated. Two doses of nitrogen (low, 50 kg ha-1 and high,
200 kg ha-1), three age categories of tillers (young, <2 months,
mature, 2 to 4 months, and old,> 4 months), and to two periods
(beginning, 1 to 45 days and end, 46 to 90) during deferral were
evaluated. The experimental design was completely randomized, in a
subsubdivide plot scheme, with four replications. In general, young tiller
had a better morphological composition (40.0% live leaf, P=0.0074; 39.2%
live stem, P=0.0208; and 20.8% dead leaf blade, P=0.0155), while the old
tiller had a worse morphology (7.3% live leaf, 54.8% live stem and 38.0%
leaf dead) at the end of the deferral period. The percentage of old
tillers was higher (P=0.0019) at the end (54.0%) than at the beginning
(43.4%) of the deferment period. At the beginning of the deferment the
canopy fertilized with high dose of N presented (P=0.0447) a higher
percentage of young tiller (36.8%) than that under low N dose (28.6%). At
the end of the deferment period, both the high and the low nitrogen doses
resulted (P=0.0276) in similar values of young tiller in the deferred
canopy (24.3%, on average). The higher N dose, plotted before the deferral
period, increases the percentage of young tiller during the initial phase
of deferment. Young tiller has better structural characteristics than
mature and old tiller.
Keywords: leaf area; morphological composition; tiller
number; Urochloa brizantha.
Recebido em 05 de dezembro de 2016.
Aceito em 07 de junho de 2019.
Introdução
Nas pastagens brasileiras ocorre a estacionalidade da produção de forragem, devido às condições climáticas variáveis ao longo do ano. Para minimizar os efeitos negativos dessa estacionalidade sobre a produção animal pode ser utilizado o diferimento das pastagens, que consiste em suspender a utilização de alguns pastos durante parte do período de maior crescimento das plantas, a fim de que a forragem produzida possa ser usada, sob pastejo, na época de escassez de alimento(1).
A adubação nitrogenada pode ser utilizada em pastagens diferidas como forma de aumentar o perfilhamento do pasto e a produção de forragem(2). Outra importante vantagem do uso do nitrogênio em pastagens diferidas é a possibilidade de redução do período de diferimento dos pastos, sem diminuir a massa de forragem diferida. A adubação nitrogenada também pode modificar a estrutura do pasto diferido. Ela corresponde à distribuição dos componentes da parte aérea da planta dentro de uma comunidade(3) e também pode ser compreendida como a morfologia do pasto que o animal encontra durante o pastejo.
Com relação ao animal, a estrutura do pasto influencia o comportamento ingestivo, o consumo e o desempenho(4). No tocante à planta forrageira, a estrutura do pasto influencia o microclima (luminosidade, umidade, temperatura, ventilação etc.) no interior do dossel, o que desencadeia modificações no desenvolvimento do pasto(5). Desse modo, a estrutura do pasto é característica determinante das respostas, tanto das plantas como dos animais, em condições de pastejo, razão de sua importância. Porém, ainda não se sabe de modo detalhado como a adubação nitrogenada modifica a estrutura do pasto diferido. Nesse sentido, o estudo das características estruturais das distintas classes de perfilhos presentes no pasto diferido pode fornecer as informações para se alcançar esse conhecimento.
O pasto diferido é composto por perfilhos com diferentes idades, que podem ser influenciados de diferentes formas pela adubação nitrogenada. De fato, em trabalhos de pesquisas com gramíneas do gênero Urochloa sob lotação contínua, tem-se demonstrado que a adubação nitrogenada aumenta a população de perfilhos jovens no pasto(6), o que pode modificar a estrutura do pasto, porque perfilhos jovens podem ter maior número de folhas e colmos mais curtos do que perfilhos velhos(7).
Assim, o objetivo com este trabalho foi compreender, por meio da avaliação das características estruturais de perfilhos com diferentes idades, como a adubação nitrogenada e a idade do perfilho modificam a estrutura do capim-marandu durante o período de diferimento.
Materiais e métodos
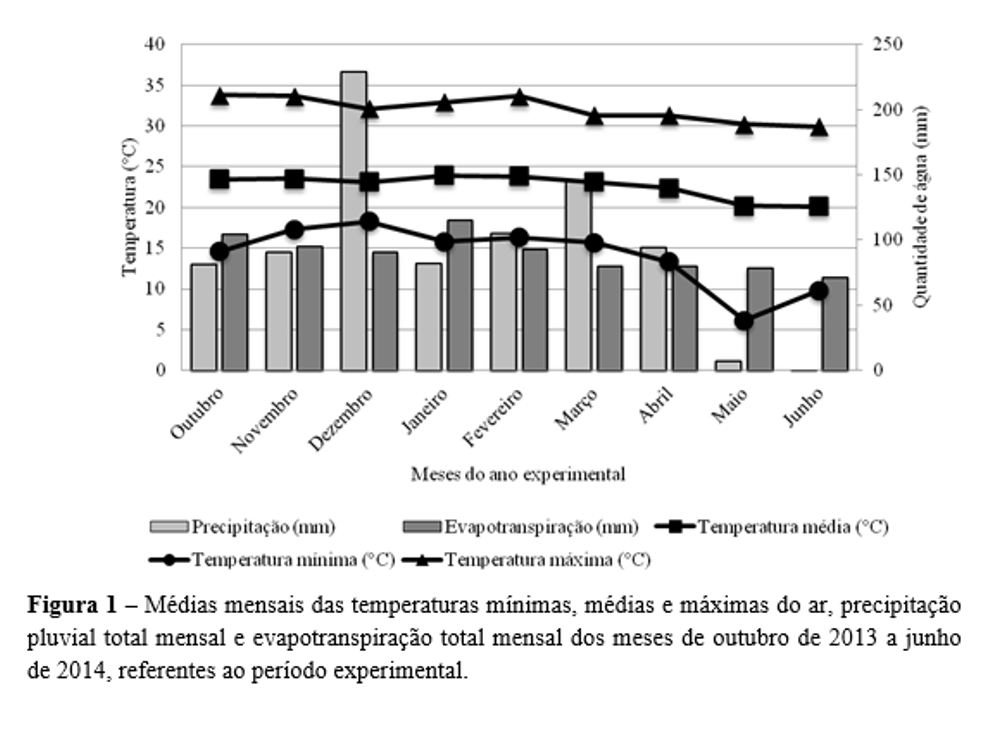
O experimento foi realizado entre outubro de 2013 e junho de 2014 na Fazenda Experimental Capim-Branco, da Universidade Federal de Uberlândia (UFU), localizada em Uberlândia, MG (18º53’19” S, 48°20’57” W e 863 metros de altitude). O pasto de Urochloa brizantha Stapf. cv. Marandu (capim-marandu) foi implantado no ano de 2000. O clima da região é Aw, tropical de savana com estação seca de inverno(8), com estação seca e chuvosa bem definida, sendo a temperatura e precipitação média anual de 22,3 ºC e 1.584 mm. Os dados climáticos durante o período experimental foram obtidos em estação meteorológica localizada a 200 m do local do experimento (Figura 1).
O solo da área experimental foi classificado como Latossolo Vermelho Escuro Distrófico e com relevo típico de chapada (relevo suavemente ondulado com formações sedimentares, apresentando vales espaçados e raros)(9). Antes da implantação do experimento foi realizada a amostragem do solo para análise química, em camada de 0 a 20 cm, que apresentou as seguintes características: pH em H2O: 5,5; P: 1,3 mg dm-3 (Mehlich-1); K: 75 mg dm-3; Ca2+: 1,7 cmolc dm-3; Mg2+: 1,1 cmolc dm-3; Al3+: 0,0 cmolc dm-3 (KCl 1 mol L-1) e P-rem: 11,7 mg dm-3. Com base nos resultados, não foi necessária realizar a calagem. Em 10 de janeiro de 2014 foram feitas as adubações potássica e fosfatada, com aplicação de 50 kg ha-1 de K2O e P2O5, utilizando como fontes o cloreto de potássio e superfosfato simples.
De outubro de 2013 a março de 2014 o capim-marandu foi mantido a 30 cm de altura, de acordo com as recomendações de Sbrissia e Da Silva(10) para o manejo do pastejo em lotação contínua. Para isso, cortes semanais foram realizados e o excesso de forragem retirado do interior das unidades experimentais.
Foram avaliadas duas estratégias de adubação nitrogenada, sendo uma dose baixa, 50 kg ha-1 de nitrogênio (N) aplicada no dia 15/03/2014, e a outra correspondeu a uma dose alta, 200 kg ha-1 de N, parcelada em três aplicações: 80 kg ha-1 (aplicada em 10/01/2014), 70 kg ha-1 (aplicada em 17/02/2014) e 50 kg ha-1 (aplicada em 15/03/2014). Foi utilizada como fonte de adubo a ureia e as adubações ocorreram em cobertura ao fim da tarde. Cada estratégia de adubação foi implementada em quatro unidades experimentais, totalizando oito parcelas com área de 9 m² cada.
Três faixas etárias de perfilhos (jovem, maduro e velho) foram avaliadas. Essa categorização foi possível mediante o uso da técnica de dinâmica de perfilhamento, que quantifica o aparecimento e morte dos perfilhos em avaliações mensais. A dinâmica de perfilhamento iniciou em outubro de 2013, sendo realizada durante todo o período experimental dentro de duas áreas delimitadas por tubo de PVC de 30 cm de diâmetro, usando fios de diferentes cores para identificar cada geração de perfilhos(11). Isso permitiu identificar os números de perfilhos jovens, com menos de dois meses de idade; maduros, entre dois e quatro meses; e velhos, com mais que quatro meses(12).
Adicionalmente, o período de diferimento de 90 dias, que iniciou em 15/03/2014 e terminou em 13/06/2014, foi dividido em um período inicial (primeiros 45 dias) e outro final (últimos 45 dias). Durante esses dois períodos a morfogênese foi avaliada em quatro perfilhos de cada faixa etária(13). Os perfilhos foram marcados nas unidades experimentais com presilhas identificadas e, em cada ciclo de avaliação, novo grupo de perfilhos foi selecionado para avaliação. Nestes perfilhos foram medidos os comprimentos do colmo e da lâmina foliar, bem como contabilizados os números de folhas vivas e mortas. Essas mensurações foram realizadas uma vez por semana. O comprimento do colmo foi mensurado desde o nível do solo até a lígula da folha mais velha, completamente expandida. O comprimento da lâmina foliar correspondeu à distância do ápice das folhas expandidas até a sua lígula. As folhas em expansão e expandidas foram consideradas vivas e as que possuíam mais de 50% da lâmina foliar senescente, mortas. Com esses dados, foram calculadas as seguintes características: comprimento da lâmina foliar (CLF), comprimento final do colmo (CFC) e os números de folhas vivas (NFV) e mortas (NFM) por perfilho.
No final do período de diferimento dez perfilhos de cada categoria de idade foram colhidos por parcela e tiveram seus componentes morfológicos separados em lâmina foliar viva, lâmina foliar morta e colmo vivo. As extremidades das lâminas foliares vivas foram cortadas, de modo a se obter um segmento aproximadamente retangular, cuja área foi estimada pela multiplicação de seu comprimento pela sua largura. Todas as subamostras foram acondicionadas em sacos de papel, secadas na estufa de ventilação forçada a 65 ºC por 72 horas e pesadas após esse período. Com esses dados foi possível calcular as percentagens de lâmina foliar viva (PLFV), de lâmina foliar morta (PLFM) e de colmo vivo (PCV) de cada categoria de perfilho. O peso médio de cada faixa etária de perfilho foi calculado pelo somatório das massas de todos os seus respectivos componentes morfológicos (lâmina foliar viva, lâmina foliar morta e colmo vivo), o qual foi dividido pelo número de perfilhos colhidos, que correspondeu a 10 perfilhos de cada faixa etária. A área foliar específica (AFE) foi calculada pela divisão da área e pelo peso dos segmentos retangulares de lâmina foliares. A área do perfilho foi obtida pela multiplicação da massa de lâmina foliar viva do perfilho pela sua área foliar específica.
Os dados foram analisados em delineamento inteiramente casualizado, com esquema de parcela sub-subdividida e quatro repetições. O critério para determinação do delineamento experimental foi o relevo uniforme e a boa cobertura do solo pela gramínea na área experimental. As estratégias de adubação nitrogenada corresponderam às parcelas, as categorias de idades de perfilhos (jovem, maduro e velho) consistiram nas subparcelas e o período de avaliação (início e fim do diferimento) foi a sub-subparcela. Portanto, o modelo experimental usado foi o seguinte:
Yijk = μ + Ni + eij + Ik + (NI)ik + eijk + Pn+ (NP)in +
(IP)kn + (NIP)ikn + eikjn
Onde:
Yijkn: valor observado na sub-subparcela n, da subparcela k, da parcela j
e repetição i;
μ: média geral;
Ni: fator da parcela principal (adubação nitrogenada);
eij: erro da parcela principal;
Ik: fator da subparcela (idade do perfilho);
(NI)ik: interação N x I;
eijk: erro da subparcela;
Pn: fator da sub-subparcela (período do diferimento);
(NP)in: interação N x P;
(IP)kn: interação I x P;
(NIP)ikn: interação N x I x P;
eikjn: erro da sub-subparcela.
Para cada característica avaliada, foi realizada análise de variância e os efeitos dos fatores foram comparados pelo teste t. Todas as análises foram realizadas ao nível de significância de até 5% de probabilidade para ocorrência do erro tipo I.
Resultados
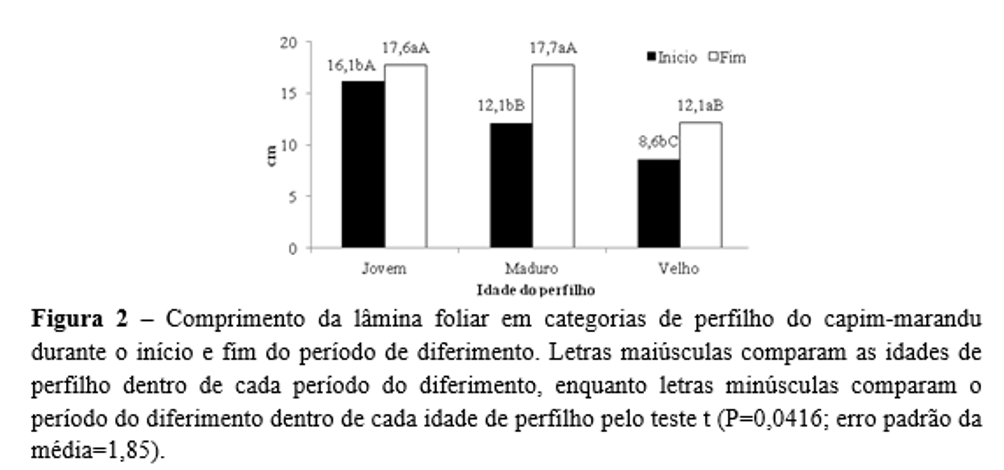
O CLF foi influenciado pela dose de nitrogênio (P=0,0307) e pela interação entre idade do perfilho e período do diferimento (P=0,0416). O CLF foi maior (P=0,0307) no capim-marandu adubado com alta (15,1 cm) do que com baixa (12,9 cm) dose de nitrogênio. No início do diferimento o CLF foi (P=0,0416) maior no perfilho jovem, intermediário no perfilho maduro e menor no perfilho velho (Figura 2). No fim do diferimento, os perfilhos jovens e maduros apresentaram (P=0,0416) valores superiores em relação ao perfilho velho (Figura 2).
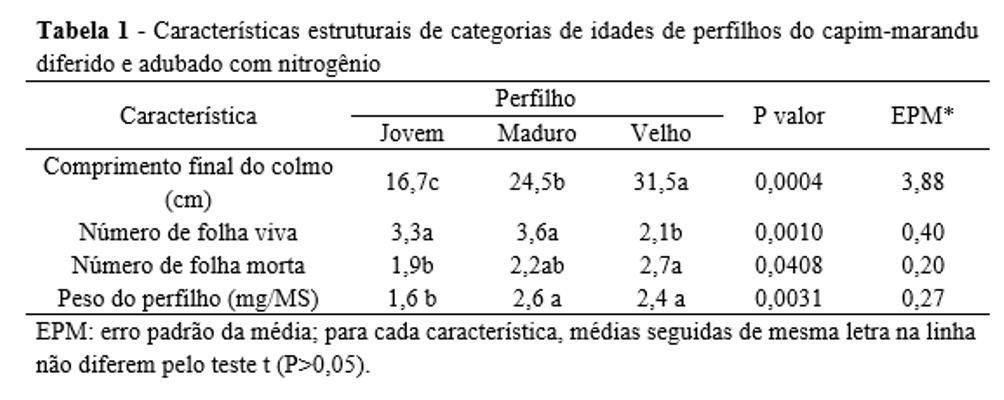
O CFC, o NFV e o NFM foram influenciados de forma isolada pela idade do perfilho e pelo período do diferimento. O CFC do capim-marandu foi (P=0,0004) maior no perfilho velho, intermediário no perfilho maduro e menor no perfilho jovem (Tabela 1). O NFV foi (P=0,0010) maior nos perfilhos jovens e maduros, em relação aos perfilhos velhos (Tabela 1). O NFM foi (P=0,0408) maior no perfilho velho e menor no perfilho jovem (Tabela 1).
O peso do perfilho foi influenciado isoladamente pela idade do perfilho (P=0,0031) e pela dose de nitrogênio (P=0,0019). O peso do perfilho foi (P=0,0031) maior nos perfilhos maduros e velhos, em relação aos perfilhos jovens (Tabela 1). O peso do perfilho ao término do período de diferimento do capim-marandu adubado com 200 kg ha-1 de N (2,5 mg) foi (P=0,0019) maior, em comparação ao capim adubado com 50 kg ha-1 de N, que apresentou peso de perfilho de 1,9 mg.
O número total de perfilho foi influenciado de forma isolada pelo período do diferimento (P=0,001) e também pela dose de nitrogênio (P=0,0092). Já a porcentagem de perfilho velho foi influenciada (P=0,0019) de forma isolada apenas pelo período do diferimento. O comprimento final do colmo (P=0,0376) e os números de folhas vivas (P<0,0001) e mortas (P<0,0001) foram maiores no início do que no fim do diferimento, um padrão de resposta contrário ao observado para o número total de perfilhos (P=0,0001) e a porcentagem de perfilho velho (P=0,0019) (Tabela 2). O número total de perfilhos também foi superior (P=0,0092) no capim-marandu adubado com alta (291 perfilhos/m²) do que com baixa (214 perfilhos/m²) dose de N.
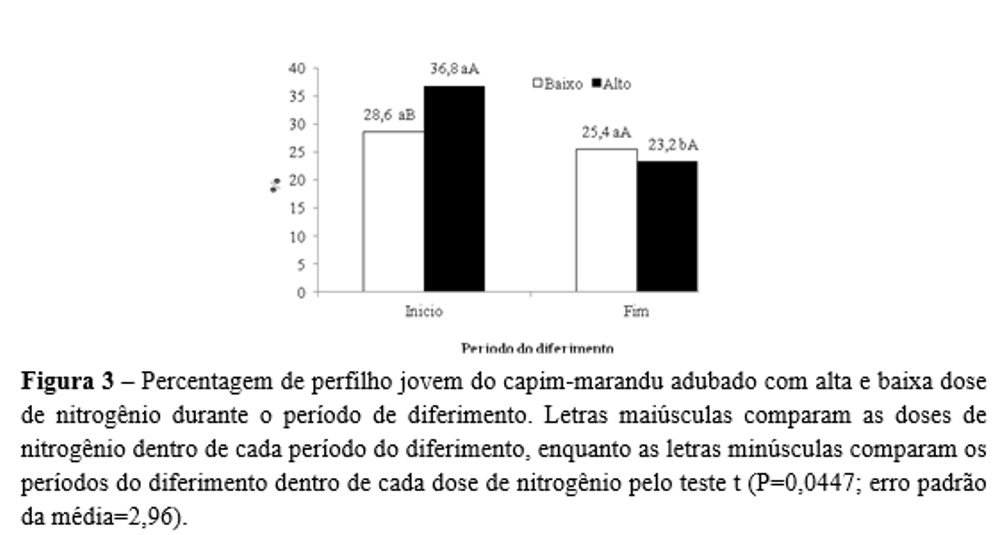
A percentagem de perfilho jovem foi influenciada (P=0,0447) apenas pela interação entre o período do diferimento e a adubação nitrogenada, sendo que, no início do diferimento, a planta adubada com maior dose de N apresentou maior valor do que aquela submetida à baixa dose de N. Em contrapartida, no fim do período de diferimento, tanto a alta como a baixa dose de nitrogênio resultaram em valores similares de percentagem de perfilho jovem. Ele foi maior no início, quando comparado ao fim do período de diferimento, quando alta dose de adubo nitrogenado foi utilizada (Figura 3).
A percentagem de perfilho maduro não foi influenciada por nenhum dos fatores estudados e apresentou valor médio de 22,6%.
A composição morfológica e a área dos perfilhos ao fim do período de diferimento foram influenciadas apenas pela interação entre idade do perfilho e dose de N (Tabela 3). A percentagem de lâmina foliar viva (PLFV) do perfilho jovem foi superior quando alta dose de N foi utilizada, contrariamente ao observado para o perfilho maduro. A PLFV do perfilho velho não foi influenciada pela dose de N (Tabela 3).
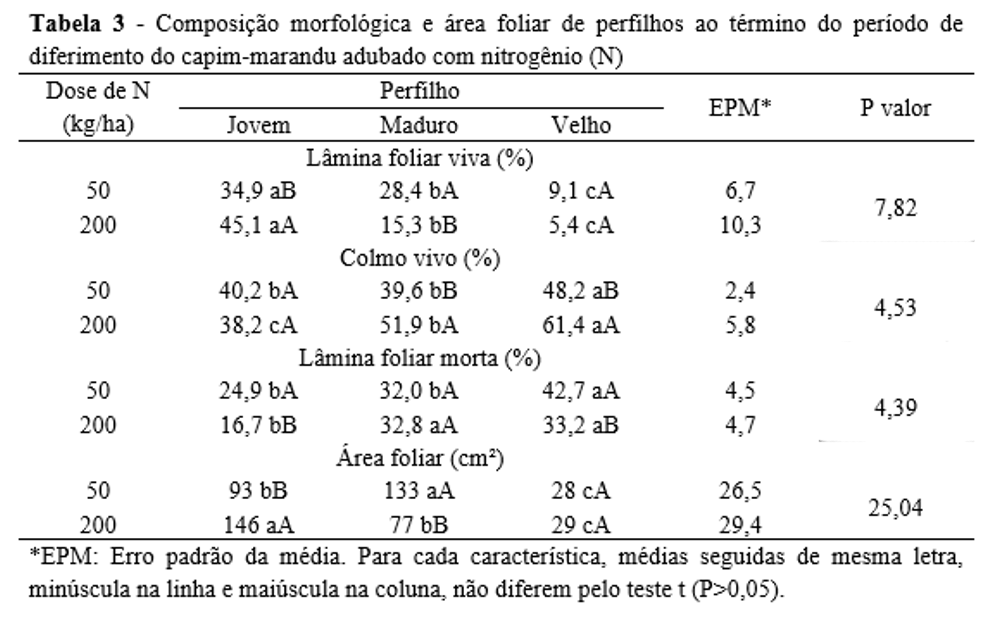
A porcentagem de colmo vivo (PCV) dos perfilhos maduros e velhos foi maior com a utilização de alta dose de N, mas essa característica não variou no perfilho jovem (Tabela 3). Em ambas as doses de N avaliadas o perfilho jovem apresentou inferior PCV em relação ao perfilho velho, um padrão de resposta semelhante ao verificado para a percentagem de lâmina foliar morta (PLFM). A PLFM do perfilho maduro não foi influenciada pela adubação nitrogenada, contudo, nos perfilhos jovens e velhos, seus valores foram superiores na condição de baixa dose de N (Tabela 3).
Com relação à área foliar, no dossel sob baixa dose de N, seus valores foram maiores no perfilho maduro e inferiores no perfilho velho. Com uso de alta dose de N, o perfilho velho também apresentou inferior área foliar, mas o maior valor foi verificado no perfilho jovem. A área foliar do perfilho velho não foi influenciada pela dose de N, porém, no perfilho jovem, maior área foliar ocorreu com a alta dose de N, contrariamente ao observado para o perfilho maduro (Tabela 3).
Discussão
O perfilho velho está em estágio de desenvolvimento mais avançado e é mais pesado, o que exige um órgão estrutural para manter sua adequada sustentação, que é o colmo(14). Também é possível que muitos perfilhos velhos tenham passado do estágio vegetativo para o reprodutivo. Com isso, há a elevação do meristema apical e, consequentemente, a distância até o ápice do perfilho reduz, o que pode ter diminuído o comprimento da lâmina foliar (CLF) do perfilho velho(15). Além disso, o perfilho velho geralmente apresenta menor taxa de alongamento foliar do que o perfilho jovem(16), o que também justifica seu inferior CLF.
Como o perfilho velho apresentou lâmina foliar mais curta, é possível que mais rapidamente a senescência tenha ultrapassado 50% do CLF, permitindo classificar a folha como morta. Adicionalmente, como o perfilho velho está em estágio de desenvolvimento mais avançado, ele pode ter apresentado menor taxa de aparecimento foliar(16), o que também justificaria seu menor NFV.
No início do diferimento, os perfilhos marcados passaram por um período com condição mais favorável ao crescimento vegetal, caracterizada por temperatura e precipitação pluvial mais altas, o que justifica seus maiores comprimentos de colmo no início da avaliação morfogênica. Com a troca dos perfilhos após os primeiros 45 dias de avaliação, o tamanho inicial do colmo dos novos perfilhos foi menor, porque eles se desenvolveram sob condições climáticas mais limitantes ao crescimento da planta. Com isso, no fim do diferimento, o comprimento do colmo reduziu.
Considerando-se que a taxa de aparecimento foliar é mais alta em condições de clima mais favorável(17), possivelmente o número de folha viva por perfilho foi maior no início do que no fim do diferimento, em razão do maior aparecimento foliar no início desse período, que apresentou características climáticas mais predisponentes ao crescimento vegetal. Com relação ao número de folha morta por perfilho, o seu valor inferior no fim do diferimento pode ter sido consequência do aumento da duração de vida da folha, típico em condições climáticas mais adversas ao vegetal(17), tais como baixa temperatura e pouca precipitação pluvial, que ocorreram ao fim do diferimento.
O número total de perfilho foi maior no fim do que no início do diferimento provavelmente porque a maioria dos perfilhos que apareceu no início do diferimento se manteve viva até o fim do diferimento e, principalmente, porque, durante o período de diferimento, novos perfilhos surgiram no dossel forrageiro.
Com relação aos efeitos do nitrogênio, o maior comprimento da lâmina foliar do capim-marandu sob alta dose de N ocorreu em função do estímulo desse nutriente sobre o alongamento foliar de gramíneas forrageiras tropicais, ao estimular a produção de células(18, 19 e 20).
Tanto o número total quanto o peso do perfilho foram maiores no dossel forrageiro sob alta dose de N, pois esse nutriente estimula concomitantemente as gemas basais a se desenvolverem em perfilhos(21) e também aumenta o crescimento de folhas e do colmo em perfilhos individuais(14, 18 e 22), desde que o índice de área foliar do pasto não seja demasiadamente elevado e capaz de desencadear alta competição por luz entre os perfilhos(23).
O efeito positivo de N sobre a percentagem de perfilho jovem no dossel diferido foi restrito apenas ao início do diferimento, uma consequência do clima mais favorável ao crescimento vegetal no início, em comparação ao fim do diferimento. Além disso, no início do diferimento, havia mais luz incidindo no interior do dossel, pois ele ainda estava em fase inicial de rebrotação. Essa é uma condição mais favorável ao aparecimento de novos perfilhos, contrariamente ao observado no fim do diferimento, quando o avanço no estágio de desenvolvimento do dossel causou maior sombreamento na base das plantas, o que inibe o perfilhamento(10).
Embora maior no fim do diferimento (Tabela 2), a ocorrência de perfilho velho (em média de 48,7%) foi alta no capim-marandu diferido. Esse resultado é típico de épocas do ano cujo clima é restritivo ao crescimento vegetal, tal como o outono, época em que ocorreu o período de diferimento. Nesta situação, a planta usa a estratégia de conservação de recursos, por meio do aumento na duração de vida das folhas e dos perfilhos(14) que, assim, ficam mais velhos.
De modo geral, o perfilho jovem apresentou melhor composição morfológica (mais lâmina foliar viva e menos colmo e lâmina foliar morta), enquanto o perfilho velho teve uma pior morfologia ao término do período de diferimento. O perfilho maduro apresentou características morfológicas intermediárias à dos demais perfilhos. Com o aumento do estádio de desenvolvimento, é natural que o perfilho fique mais pesado e, com efeito, o colmo torna-se mais desenvolvido(14), a fim de sustentar o maior peso da planta. Com o desenvolvimento do perfilho, algumas de suas folhas ultrapassam o tempo em que permanecem vivas(24), o que aumenta a percentagem de lâmina foliar morta no perfilho maduro e, sobretudo, no perfilho velho.
A adubação nitrogenada aumenta o crescimento foliar do perfilho jovem(7), o que justifica a maior percentagem de lâmina foliar viva nessa classe de perfilho com a aplicação de maior dose de N. O elevado desenvolvimento do colmo em perfilhos maduros e velhos sob alta dose de N, além de aumentar o percentual de colmo na massa do perfilho, também fez com que a percentagem de lâmina foliar viva fosse reduzida no perfilho maduro e ficasse inalterada no perfilho velho.
Os perfilhos maduros e velhos, por serem mais desenvolvidos, tendem a alongar mais o colmo, o que parece ter sido estimulado pela aplicação de alta dose de N, que proporcionou maiores percentuais de colmo nessas classes de perfilhos. Por outro lado, a percentagem de colmo do perfilho jovem não foi influenciada pela aplicação de N, provavelmente porque essa classe de perfilhos tem o colmo naturalmente mais curto.
O maior estádio de desenvolvimento do perfilho velho justifica seu maior percentual de lâmina foliar morta ao término do diferimento no dossel sob baixa dose de N. Adicionalmente, é possível que a alta disponibilidade de N tenha aumentado a taxa de desenvolvimento do perfilho maduro que, assim, apresentou diferenciação morfológica semelhante à observada no perfilho velho, razão pela qual, no dossel adubado com alta dose de N, além do perfilho velho, o maduro também apresentou alto percentual de lâmina foliar morta em relação ao perfilho jovem.
É possível que o perfilho jovem ainda não tivesse alcançado o seu número potencial de folha viva, enquanto o perfilho maduro pode ter apresentado maior senescência foliar, razão pela qual as áreas foliares desses perfilhos foram inferiores à do perfilho maduro no dossel sob aplicação de baixa dose de N. Por outro lado, conforme já relatado, é provável que, sob alta dose de N, a taxa de desenvolvimento dos perfilhos tenha aumentado, o que pode ter antecipado a ocorrência de senescência foliar naqueles perfilhos de maior idade. Com isso, a área foliar reduziu gradativamente com a idade do perfilho.
Com base nos dados apresentados neste trabalho, constata-se que a idade é fator determinante das características morfológicas dos perfilhos presentes no pasto diferido, de tal forma que os perfilhos mais jovens possuem melhores características estruturais do que os perfilhos mais velhos. Desta forma, estratégias de manejo idealizadas para reduzir o perfil de idade dos perfilhos no pasto diferido contribuem para a melhoria da estrutura do pasto, com prováveis consequências positivas sobre o consumo e desempenho animal em pasto. Nesse sentido, dentre as estratégias de manejo capazes de aumentar a participação relativa de perfilho jovem no pasto diferido, destacam-se a adubação nitrogenada, concomitante à redução do período de diferimento.
Conclusões
Em dosséis diferidos de Urochloa brizantha cv. Marandu, a maior dose de nitrogênio, parcelada antes do período de diferimento, resulta em aumento do percentual de perfilho jovem durante a fase inicial do diferimento. O perfilho jovem tem melhores características estruturais do que os perfilhos maduros e velhos, porém o perfilho velho tem maior participação relativa no dossel diferido.
Referências
1. Santos MER, Fonseca DM, Euclides VPB, Nascimento Júnior
D, Queiróz AC, Ribeiro Júnior JI. Características estruturais e índice de
tombamento de Brachiaria decumbens cv. Basilisk em pastagens
diferidas. Revista Brasileira de Zootecnia. 2009; (38)4:626-634.
Disponível em: http://www.scielo.br/pdf/rbz/v38n4/06.pdf.
2. Sousa BML, Vilela HH, Santos AL, Santos MER, Nascimento Júnior D, Assis
CZ, Faria BD, Rocha GO. Piata palisadegrass deferred in the fall: effects
of initial height and nitrogen in the sward structure. Revista Brasileira
de Zootecnia. 2012; (41)5:1134-1139. Disponível em: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1516-35982012000500008.
3. Laca EA, Lemaire G. Measuring sward structure. In: Mannetje L, Jones
RM. Field and laboratory methods for grassland and animal production
research. Wallingford: CAB International. 2000;103-122.
4. Carvalho PCF. Harry Stobbs Memorial Lecture: Can grazing behavior
support innovations in grassland management? Tropical Grasslands. 2013;
1:137–155. Disponível em: http://www.tropicalgrasslands.info/index.php/tgft/article/view/81/44.
5. Chapman D, Lemaire G. Tissue flows in grazed plant communities. In:
Hodgson J, Illius AW. The ecology and management of grazing systems.
Guilford: CAB International. 1996; 3-36.
6. Morais RV, Fonseca DM, Nascimento Júnior D, Ribeiro Júnior JI, Fagundes
JL, Moreira LM, Mistura C, Martuscello, JA. Demografia de perfilhos
basilares em pastagens de Brachiaria decumbens adubada com
nitrogênio. Revista Brasileira de Zootecnia. 2006; (35):380–388. Diponível
em: http://www.scielo.br/pdf/rbz/v35n2/a07v35n2.pdf.
7. Paiva AJ, Da Silva SC, Pereira LET, Guarda VD, Mesquita P, Caminha FO.
Structural characteristics of tiller age categories of continuously
stocked marandu palisade grass swards fertilized with nitrogen. Revista
Brasileira de Zootecnia. 2013; (41):24-29. Disponível em: http://www.scielo.br/pdf/rbz/v41n1/04.pdf.
8. Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G. Köppen’s
climate classification map for Brazil. Meteorologische Zeitschrift. 2013;
22(6):711–728. Disponível em: http://dx.doi.org/10.1127//0941-2948/2013/0507.
9. EMBRAPA. Sistema brasileiro de classificação de solos. 2nd ed. Rio de
Janeiro: Embrapa Solos; 2006; 306p.
10. Sbrissia AF, Silva SC. Compensação tamanho/densidade populacional de
perfilhos em pastos de capim-marandu. Revista Brasileira de Zootecnia.
2008; 37(1):35-47. Disponível em: http://dx.doi.org/10.1590/S1516-35982008000100005.
11. Sousa BML, Santos MER, Vilela HH, Silveira MCT, Rocha GO, Freitas CAS,
Silva NAM, Nascimento Júnior, D. Piata palisade grass deferred with two
distinct initial heights: luminous environment and tillering dynamics.
Revista Brasileira de Zootecnia. 2013; 42(1):36-43. Disponível em: http://ref.scielo.org/nsxx7g.
12. Paiva AJ, Pereira LET, Silva SC, Dias RAP. Identification of tiller
age categories 17 based on morphogenetic responses of continuously stocked
marandu palisade grass fertilised with nitrogen. Ciência Rural. 2015;
45(5):867-870. Disponível em: http://dx.doi.org/10.1590/0103-8478cr20120738.
13. Barbosa RA, Nascimento Júnior D, Vilela HH, Sousa BML, Da SILVA SC,
Euclides VPB, Silveira MCT. Morphogenetic and structural characteristics
of guinea grass tillers at different ages under intermittent stocking.
Revista Brasileira de Zootecnia. 2012; (41):1583-1588. Disponível em: http://www.scielo.br/pdf/rbz/v41n7/05.pdf.
14. Santos MER, Fonseca DM, Balbino EM, Monnerat JPIS, Silva SP.
Capim-braquiária diferido e adubado com nitrogênio: produção e
características da forragem. Revista Brasileira de Zootecnia. 2009;
38(4):650-656. Disponível em: http://dx.doi.org/10.1590/S1516-35982009000400009.
15. Skinner RH, Nelson CJ. Elongation of the grass leaf and its
relationship to the phyllochron. Crop Science. 1995; (35)1:4-10.
16. Paiva AJ, Silva SC, Pereira LET, Caminha FO, Pereira PM, Guarda V.
Morphogenesis on age categories of tillers in marandu palisadegrass.
Scientia Agricola. 2011; (68)6:626-6 631. Disponível em: http://www.scielo.br/pdf/sa/v68n6/v68n6a03.pdf.
17. Santos MER, Fonseca DM, Gomes VM, Gomide CAM, Nascimento Júnior D,
Queiroz DS. Capim-braquiária sob lotação contínua e com altura única ou
variável durante as estações do ano: morfogênese e dinâmica de tecidos.
Revista Brasileira de Zootecnia. 2011; (40)11:2323-2331 Disponível em: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1516-35982011001100007.
18. Paciullo DSC, Fernandes PB, Gomide CAM, Castro CRT, Sobrinho FS,
Carvalho CAB. The growth dynamics in Brachiaria species according to
nitrogen dose and shade. Revista Brasileira de Zootecnia. 2011;
(40)2:270-276. Disponível em: http://www.scielo.br/pdf/rbz/v40n2/06.pdf.
19. Garcez Neto AF, Nascimento Júnior D, Regazzi AJ, Fonseca DM, Mosquim
PR, Gobbi KF. Respostas morfogênicas e Estruturais de Panicum maximum
cv. Mombaça sob diferentes níveis de adubação nitrogenada e alturas de
corte. Revista Brasileira de Zootecnia. 2002; (31)5:1890-1900. Disponível
em: http://www.scielo.br/pdf/rbz/v31n5/a04v31n5.pdf.
20. Martuscello JA, Fonseca DM, Nascimento Júnior D, Santos PM, Ribeiro
Júnior JI, Cunha DNFV, Moreira LM. Características morfogênicas e
estruturais do capim-xaraés submetido à adubação nitrogenada e
desfolhação. Revista Brasileira de Zootecnia. 2005; (34)5: 1475-1482.
Disponível em: http://www.scielo.br/pdf/rbz/v34n5/26627.pdf.
21. Iwamoto BS, Cecato U, Ribeiro OL, Mari GC, Peluso EP, Lourenço DAL.
Características morfogênicas do capim-tanzânia fertilizado ou não com
nitrogênio nas estações do ano. Bioscience Journal. 2015;(31)1: 181-193.
22. Fagundes JL, Fonseca DM, Mistura C, Morais RV, Vitor CMT, Gomide JA,
Nascimento Júnior D, Casagrande DR, Costa LT. Características morfogênicas
e estruturais do capim-braquiária em pastagem adubada com nitrogênio
avaliadas nas quatro estações do ano. Revista Brasileira de Zootecnia.
2006;(35)1:21-29. Disponível em: http://www.scielo.br/pdf/rbz/v35n1/28338.pdf.
23. Matthew C, Assuero SG, Black CK, Sackville Hamilton NR. Tiller
dynamics of grazed swards. In: Lemaire G, Hodgson J, Moraes A, Nabinger C,
Carvalho PCR. Grassland ecophysiology and grazing ecology. Wallingford:
CAB International. 2000; 127-150.
24. Santos MER, Fonseca DM, Braz TGS, Silva SP, Gomes VM, Silva GP.
Características morfogênicas e estruturais de perfilhos de
capim-braquiária em locais do pasto com alturas variáveis. Revista
Brasileira de Zootecnia. 2011; (40)3:535-542. Disponível em: http://www.scielo.br/pdf/rbz/v40n3/10.pdf.